


Effect of Different Maceration Times on the Composition of Condensed Tannins and Sensory Quality of Wines
-
摘要: 为明确不同皮渣浸渍时间对葡萄酒中缩合单宁的影响,以赤霞珠和西拉为材料,进行不同浸渍发酵时间处理,对葡萄皮和葡萄籽以及葡萄酒中缩合单宁的含量、组成、缩合单宁以及相关口感指标等进行了分析。结果表明,不同葡萄品种中缩合单宁的初始含量、聚合度及没食子酰化率都有显著差异(P<0.05)。葡萄皮中缩合单宁的平均聚合度显著高于葡萄籽(P<0.05),而葡萄皮缩合单宁的没食子酰化率则显著低于葡萄籽缩合单宁(P<0.05)。葡萄皮和葡萄籽中缩合单宁组成的最大差异是葡萄籽缩合单宁中不含有(−)-表棓儿茶素亚单元,而且末端单元主要是由(+)-儿茶素、(−)-表儿茶素和(−)-表儿茶素没食子酸酯组成。皮渣浸渍时间对葡萄酒中缩合单宁的组成及含量都有明显影响。随着浸渍时间的延长,葡萄酒中缩合单宁的总含量呈上升趋势,而平均聚合度则呈下降趋势。不同品种的葡萄酒之间缩合单宁含量也存在显著差异(P<0.05),这与不同葡萄品种中酚类物质的可浸提率有关。葡萄酒中葡萄皮缩合单宁的占比一直显著高于籽粒缩合单宁的占比(P<0.05),说明葡萄皮中缩合单宁比葡萄籽缩合单宁更容易浸提到葡萄酒中。相对于苦味和酸味,不同浸渍时间处理对葡萄酒的涩味影响更为显著(P<0.05),综合考虑酒体的丰满度和口感的平衡,皮渣浸渍时间选择14 d左右更好一些。Abstract: In order to study the effects of different maceration times on the condensed tannin profiles of wine, the content, composition of condensed tannins and mouth-feel attributes related with condensed tannins in grape berries and resulting wines were analyzed with Cabernet Sauvignon and Syrah as materials. The results showed that the initial concentration, degree of polymerization, extent of galloylation of condensed tannins in different grape varieties were different significantly (P<0.05). The mean degree of polymerization of condensed tannins in grape skin was significantly higher than that in grape seed (P<0.05), while the extent of galloylation of condensed tannins in skin was lower than that in seed (P<0.05). The major difference between the composition of condensed tannins in grape skin and seed was the absence of (−)-epigallocatechin units, with (+)-catechin, (−)-epicatechin and (−)-epicatechin-3-O-gallate as terminal units. The different maceration times affected the profile of condensed tannins in resulting wines. With the prolongation of maceration time, the total content of condensed tannin in wine tended to increase, while the polymerization degree decreased. There was also significant difference in condensed tannin content among different grape varieties (P<0.05), which was related to the extraction rate of phenols in different grape varieties. The percentages of skin-derived condensed tannins were always higher than that of seed-derived condensed tannins in two variety wines (P<0.05), suggesting that skin condensed tannins were more readily extracted than seed condensed tannins. The maceration time affected the astringency more significant than bitterness and acidity (P<0.05). Considering the fullness of the wine body and the balance of taste, 14 days of the maceration time was recommended.
-
Keywords:
- condensed tannin /
- pomace /
- maceration time /
- grape /
- wine
-
缩合单宁(Condensed tannins)又称原花色素(Proanthocyanidins,PAs),主要是以黄烷-3-醇为基本单元,通过C4-C8或C4-C6共价键连接而成的多聚体,其中根据黄烷-3-醇单元B环羟基数目的不同和C环3-羟基结构的不同,主要可分为儿茶素(CAT)、表儿茶素(EC)、表没食子儿茶素(EGC)和表儿茶素没食子酸酯(ECG)等,其酚羟基基团能与蛋白质、金属离子、花色苷、多糖等结合[1-3]。作为葡萄与葡萄酒中一类重要的多酚物质,其含量、结构、聚合度、没食子酰化度等对葡萄酒感官特性,如色泽、澄清度、酒体、苦涩味等以及稳定性、陈酿潜力等都具有直接或间接的影响[4-5]。
葡萄酒中的缩合单宁主要来源于葡萄皮和种子,发酵过程中通过皮渣浸渍作用等进入葡萄酒中。葡萄原料中的缩合单宁的含量、聚合度、结构等会因葡萄品种、不同组织部位、葡萄成熟度等而有差异,如葡萄皮缩合单宁中含有原花青靛(Prodelphinidins,PDs),而种子中则没有;葡萄皮缩合单宁的平均聚合度(mDP)一般为20,而种子中的平均聚合度一般为5~8;葡萄皮中缩合单宁构成单元主要由CAT、EC、ECG、EGC等组成,而葡萄种子中则主要由CAT、EC、ECG等组成[6-7]。研究人员曾利用不同的分析方法对不同品种葡萄果实中的缩合单宁进行过测定,研究结果表明,葡萄收获时果实中的缩合单宁含量并不能代表最终能浸提到葡萄酒中的缩合单宁水平[8-9],因为除了葡萄原料之外,酿造过程中发酵温度、皮渣浸渍时间、低温预发酵工艺、皮汁比调整、浸渍酶添加等不同的酿酒工艺,都会影响成品葡萄酒中酚类物质的含量以及组成[10-11],研究酿造工艺对最终葡萄酒产品中缩合单宁的组成和含量具有重要的意义。基于此,本文主要通过探究葡萄皮渣浸渍时间对葡萄酒中缩合单宁的组成以及感官品质的影响,以期对葡萄酒酿造工艺的改进有所助益。
1. 材料与方法
1.1 材料与仪器
酿酒葡萄赤霞珠、西拉 于2017年4月采自新西兰林肯大学葡萄种质资源圃,每个品种选取20株葡萄,每株葡萄随机选择葡萄穗,在葡萄穗的不同部位摘取5~10粒葡萄,共摘取大约600 g葡萄粒,分成3份保存;酵母Lalvin D254 法国拉曼公司;亚硫酸 帝伯仕公司;丙酮、甲醇、乙腈、甲酸、乙酸 HPLC 色谱级,美国Fisher公司;(+)-儿茶素、(−)-表儿茶素、(−)-表棓儿茶素、(−)-表棓儿茶素没食子酸酯、三氟乙酸、抗坏血酸、醋酸钠 美国Sigma-Aldrich公司;Toyopearl HW-50(F) 日本Tosoh公司;实验用水 超纯水(电导度18 M)。
Waters 2695高效液相色谱仪、Waters 2996光电二极管阵列检测器(PDA) 美国Waters公司;Multifuge X1R台式高速冷冻离心机 美国Thermo Fisher公司;Eyela N 1100旋转蒸发仪 日本Eyela公司;Labconco Centrivap真空离心浓缩仪 美国Labconco公司;Christ Alpha 1-4 LSC plus冷冻干燥机 德国Christ公司。
1.2 实验方法
1.2.1 不同皮渣浸渍发酵工艺
参考李华等[12]的工艺流程。酿酒葡萄→除梗破碎→葡萄浆→添加亚硫酸(60 mg/L)→添加酵母(200 mg/L)→ 25 ℃浸渍发酵(皮渣浸渍7、14、21 d,期间每天压帽2~3次)→皮渣分离(1.5 bar压榨)→自流汁与压榨汁混合进入后酵(25 ± 1 ℃)→待还原糖降至2 g/L时发酵结束→倒罐→下胶澄清(皂土,添加量通过试验确定,参考用量0.3~0.6 g/L)→纸板过滤→充氮气低温储存(12~15 ℃)。
1.2.2 葡萄皮和籽中缩合单宁的分离
参考Kennedy等[13]的方法,并稍作修改。取10粒葡萄,称重,手动分离葡萄皮和种子,用双蒸水冲洗。分别放入具塞离心管中,加入1 mL/g粒重的丙酮:水(3:2,v/v)提取液,室温避光振荡提取24 h。提取液低温离心15 min(8000 r/min,4 ℃),上清液进行真空旋蒸,冻干成粉。用1 mL甲醇重溶,置于2 mL棕色样品瓶中,−20 ℃冷冻保存备用。所有取样和分析均3次重复。
间苯三酚酸催化降解反应:取100 μL甲醇重溶液,加入100 μL间苯三酚反应试剂,50 ℃水浴反应20 min后,加入400 μL 0.2 mol/L醋酸钠溶液,终止反应。间苯三酚反应试剂的配制:0.2 mol/L盐酸甲醇溶液,含100 g/L间苯三酚和20 g/L抗坏血酸。
1.2.3 葡萄酒中缩合单宁的分离
参考Pastor等[14]的方法,并进行适当优化。取5 mL葡萄酒样品,真空浓缩仪浓缩,用3 mL水:乙酸(98:2,v/v)重溶,上Waters C18固相萃取小柱。样品完全吸附后,用20 mL水:乙酸(98:2,v/v)洗柱,再用10 mL甲醇洗脱,得到的甲醇洗脱液进行真空旋蒸,冻干成粉。用1 mL甲醇重溶,置于2 mL棕色样品瓶中,−20 ℃避光保存备用。间苯三酚酸催化降解反应同1.2.2。
1.2.4 缩合单宁的HPLC分析条件
HPLC分析方法参考Ducasse等[15]的方法,并稍作修改。Waters Atlantis dC18(250 mm×4.6 mm, 5 μm)色谱柱,Waters保护柱(20 mm×4.6 mm,5 μm),紫外检测波长为280 nm,柱温30 ℃,进样量为10 μL,流速为0.8 mL/min。梯度洗脱:流动相A:水:甲酸(98:2,v/v),流动相B:乙腈:流动相A(80:20,v/v)。洗脱程序:0~5 min,0 B;5~35 min,0~10% B;35~70 min,10%~20%B;70~75 min,20%~100% B;75~80 min,100%~0 B。缩合单宁构成单元根据保留时间以及参考已报道的资料文献进行定性[13],并以(+)-儿茶素(CAT)为标准进行外标法定量,相关计算参考文献[7,13]的方法。
1.2.5 感官分析
由10名品酒员组成品评小组,在正式感官评定前,先用酒石酸、硫酸奎宁、硫酸铝等配制不同浓度的模拟酒液,分别进行酸味、苦味、涩味的品评训练。然后再对不同浸渍时间处理的葡萄酒样品进行感官品评分析,主要对酒样的酸度、涩度以及苦味进行评价和描述,采取8分制,8分表示强度最高。根据定量描述分析结果绘制蛛网图。
1.3 数据处理
数据采用SPSS 18.0统计软件分析。通过邓肯检验和ANOVA方差分析法进行变量间差异显著性分析。
2. 结果与分析
2.1 不同品种葡萄原料中缩合单宁组成及含量
葡萄原料中的缩合单宁含量会直接影响葡萄酒中的浓度。从表1可以看出,赤霞珠葡萄皮中缩合单宁含量大约为9.42 mg/g皮,显著(P<0.05)高于西拉葡萄皮中的含量(5.32 mg/g皮);百克鲜重含量大约为80.92 mg/100 g,也明显高于西拉葡萄的44.36 mg/100 g。赤霞珠葡萄皮缩合单宁的聚合度mDP约为23.50,明显高于西拉葡萄皮的14.61,这与文献[16]的研究结果基本一致。此外,西拉葡萄皮中缩合单宁没食子酰化度(4.22%)要显著(P<0.05)大于赤霞珠葡萄皮(2.41%)。
从表2可以看出,赤霞珠葡萄籽中缩合单宁含量约为24.56 mg/g籽,略高于西拉葡萄籽中的含量20.89 mg/g籽;2个品种葡萄籽缩合单宁平均聚合度mDP没有明显差异。此外,西拉葡萄种子中缩合单宁的没食子酰化度(20.65%)显著(P<0.05)高于赤霞珠葡萄种子(15.82%)。
表 1 赤霞珠和西拉葡萄皮中缩合单宁的含量及组成Table 1. Contents and composition of condensed tannins in CS and Syrah skin指标 赤霞珠 西拉 缩合单宁含量CTs (mg/g皮) 9.42 ± 0.31a 5.32 ± 0.12b 缩合单宁含量CTs (mg/100 g葡萄) 80.92 ± 3.10a 44.36 ± 2.48b 平均聚合度mDP 23.50 ± 2.31a 14.61 ± 1.39b 没食子酰化率G(%) 2.41 ± 0.37b 4.22 ± 0.76a 儿茶素末端单元比tCAT(%) 3.12 ± 0.05b 6.27 ± 0.09a 表儿茶素末端单元比tEC(%) 1.72 ± 0.02b 2.70 ± 0.04a 表儿茶素没食子酸酯末端单元比tECG(%) 0.14 ± 0.01a 0.16 ± 0.01a 儿茶素延伸单元比ext CAT(%) 1.78 ± 0.03a 1.71 ± 0.03a 表儿茶素延伸单元比ext EC(%) 36.61 ± 0.16b 54.40 ± 0.25a 表儿茶素没食子酸酯延伸单元比ext ECG(%) 2.01 ± 0.06b 4.27 ± 0.08a 表棓儿茶素延伸单元比ext EGC(%) 54.61 ± 0.31a 30.50 ± 0.12b 注:同行不同小写字母表示差异显著(P<0.05);表2~表3同。 表 2 赤霞珠和西拉葡萄籽中缩合单宁的含量及组成Table 2. Content and composition of condensed tannins in CS and Syrah seed指标 赤霞珠 西拉 缩合单宁含量CTs (mg/g籽) 24.56 ± 1.41a 20.89 ± 1.38b 缩合单宁含量CTs (mg/100 g葡萄) 99.81 ± 3.54a 87.78 ± 3.38b 平均聚合度mDP 6.41 ± 0.32a 6.22 ± 0.31a 没食子酰化率G(%) 15.82 ± 0.45b 20.65 ± 0.51a 儿茶素末端单元比tCAT(%) 7.12 ± 0.08a 6.94 ± 0.11a 表儿茶素末端单元比tEC(%) 5.20 ± 0.07b 6.28 ± 0.08a 表儿茶素没食子酸酯末端单元比tECG(%) 2.11 ± 0.05b 3.57 ± 0.06a 儿茶素延伸单元比ext CAT(%) 3.89 ± 0.07b 5.93 ± 0.10a 表儿茶素延伸单元比ext EC(%) 70.48 ± 0.65a 62.67 ± 0.56b 表儿茶素没食子酸酯延伸单元比ext ECG(%) 11.20 ± 0.15b 14.61 ± 0.17a 表棓儿茶素延伸单元比ext EGC(%) − − 从表1和表2还可以看出,2个品种葡萄皮缩合单宁末端单元主要是由(+)-儿茶素(CAT)组成,其次是(−)-表儿茶素(EC);而延伸单元则主要是由(−)-表棓儿茶素(EGC)组成和(−)-表儿茶素(EC)主导组成,这与文献[8]研究结果一致。2个品种葡萄籽缩合单宁末端单元均主要由(+)-儿茶素(CAT)、(−)-表儿茶素(EC)组成,其次是(−)-表儿茶素没食子酸酯(ECG),而延伸单元则主要由(−)-表儿茶素(EC)组成,其次是(−)-表儿茶素没食子酸酯(ECG),不含有(−)-表棓儿茶素(EGC)延伸单元,这与文献[13]研究结果一致。从表1与表2中还可以看出,葡萄籽缩合单宁含量显著(P<0.05)高于葡萄皮中的含量;葡萄皮缩合单宁的平均聚合度mDP显著(P<0.05)高于葡萄籽,而其没食子酰化度则显著(P<0.05)低于葡萄籽缩合单宁。
2.2 皮渣浸渍时间对缩合单宁的影响
从表3中可以看出,皮渣浸渍时间对不同品种葡萄酒中缩合单宁的组成及含量都有明显影响,随着浸渍时间的延长,缩合单宁的总含量呈上升趋势。
表 3 不同浸渍时间处理赤霞珠和西拉葡萄酒中缩合单宁组成及含量Table 3. Content and composition of condensed tannins in CS and Syrah wines produced with different maceration times指标 赤霞珠 西拉 7 d 14 d 21 d 7 d 14 d 21 d 缩合单宁含量CTs (mg/L) 164.36 ± 10.89c 369.74 ± 19.23b 408.05 ± 22.45a 340.98 ± 18.98c 513.90 ± 28.86b 565.14 ± 29.98a 平均聚合度mDP 8.43 ± 0.22a 7.47 ± 0.20b 6.87 ± 0.18c 7.84 ± 0.21a 7.13 ± 0.18b 6.24 ± 0.16c 没食子酰化率G(%) 1.79 ± 0.12b 1.91 ± 0.12b 2.65 ± 0.15a 3.03 ± 0.18b 3.69 ± 0.16a 3.82 ± 0.18a 儿茶素末端单元比tCAT(%) 12.40 ± 0.22a 11.01 ± 0.20b 12.89 ± 0.25a 10.14 ± 0.18c 11.12 ± 0.20b 12.98 ± 0.23a 表儿茶素末端单元比tEC(%) 2.55 ± 0.03b 1.86 ± 0.01c 3.25 ± 0.02a 3.32 ± 0.03b 3.09 ± 0.02b 4.32 ± 0.04a 表儿茶素没食子酸酯末端单元比tECG(%) 0.18 ± 0.01a 0.16 ± 0.01b 0.15 ± 0.01b 0.31 ± 0.02a 0.34 ± 0.02a 0.26 ± 0.01b 儿茶素延伸单元比ext CAT(%) 6.98 ± 0.12a 4.07 ± 0.10b 3.24 ± 0.10b 1.05 ± 0.06b 1.32 ± 0.06b 1.81 ± 0.08a 表儿茶素延伸单元比ext EC(%) 39.89 ± 0.16c 42.38 ± 0.20b 44.32 ± 0.21a 50.46 ± 0.25a 50.30 ± 0.23a 49.67 ± 0.20b 表棓儿茶素延伸单元比ext EGC(%) 36.97 ± 0.19b 39.26 ± 0.24a 34.45 ± 0.22b 32.49 ± 0.20a 31.31 ± 0.22a 28.15 ± 0.16b 表儿茶素没食子酸酯延伸单元比ext ECG(%) 1.02 ± 0.01b 1.26 ± 0.02b 1.69 ± 0.02a 2.23 ± 0.06c 2.52 ± 0.08b 2.78 ± 0.07a 葡萄皮衍生缩合单宁含量(mg/L) 126.98 ± 6.56b 267.66 ± 11.22a 283.84 ± 12.23a 283.91 ± 12.89b 419.03 ± 14.56a 431.31 ± 16.87a 酒中葡萄皮衍生缩合单宁占比(%) 77.26 ± 0.32a 72.39 ± 0.28b 69.56 ± 0.20c 83.26 ± 0.27a 81.54 ± 0.26a 76.32 ± 0.22b 葡萄籽衍生缩合单宁含量(mg/L) 59.90 ± 2.56c 89.06 ± 3.10b 147.41 ± 5.23a 43.21 ± 1.89c 82.38 ± 2.89b 112.10 ± 4.01a 酒中葡萄籽衍生缩合单宁占比(%) 22.74 ± 0.18c 27.61 ± 0.21b 30.44 ± 0.22a 16.74 ± 0.12b 18.46 ± 0.14b 23.68 ± 0.20a 葡萄皮缩合单宁浸提率(%) 10.19 ± 0.09b 21.50 ± 0.12a 22.78 ± 0.12a 41.60 ± 0.29b 61.37 ± 0.28a 63.19 ± 0.34a 葡萄籽缩合单宁浸提率(%) 3.90 ± 0.10c 5.79 ± 0.13b 9.58 ± 0.16a 3.19 ± 0.10c 6.11 ± 0.12b 8.32 ± 0.17a 由表1~表3可得,西拉葡萄原料中的葡萄皮缩合单宁含量(443.6 mg/kg葡萄)显著(P
<0.05)低于赤霞珠葡萄皮缩合单宁含量(809.2 mg/kg葡萄)(表1),而西拉葡萄籽缩合单宁含量(877.8 mg/kg葡萄)也略低于赤霞珠葡萄籽缩合单宁含量(998.1 mg/kg葡萄)(表2),但是对葡萄酒的检测结果却显示,3个处理的西拉葡萄酒中的缩合单宁含量都比相应处理的赤霞珠葡萄酒中的含量高,这可能与不同葡萄品种的葡萄皮细胞壁的成分和组成有关系,不同品种葡萄皮细胞壁中成分的组成和含量不同会影响酚类物质的浸提率[17-19],因此原料中酚类物质含量高的葡萄品种,其酒中的多酚含量不一定高,这可能是葡萄酒中缩合单宁含量与葡萄原料中的缩合单宁含量不同的原因。 从表3中还可以看出,随着浸渍时间的延长,2个品种葡萄酒中缩合单宁平均聚合度mDP均呈明显下降趋势,这可能是由于浸渍初期缩合单宁主要是来自于葡萄皮,而随着浸渍时间的延长,葡萄籽缩合单宁的浸提量开始增加,从而使得平均聚合度mDP相应出现下降[6-8],这一趋势与葡萄酒中葡萄皮和葡萄籽缩合单宁的含量变化趋势相符合。从表3中看出,葡萄酒中葡萄皮缩合单宁的含量随着初期浸渍时间的延长有所上升,但到后期浸渍14和21 d的含量则变化不再显著,而酒中葡萄籽缩合单宁的含量则随着浸渍时间的延长而呈显著(P<0.05)上升趋势。
如表1和表2所示,葡萄原料中葡萄皮缩合单宁含量显著(P<0.05)低于葡萄籽缩合单宁含量,但从表3来看,葡萄酒中葡萄皮缩合单宁的占比却一直显著(P<0.05)高于种子缩合单宁的占比,而且赤霞珠和西拉葡萄中葡萄皮缩合单宁的浸提率分别为10.19%~22.78%、41.60%~63.19%,而葡萄籽缩合单宁的浸提率则分别为3.90%~9.58%、3.19%~8.32%,这可能是由于葡萄皮中缩合单宁比葡萄籽缩合单宁更容易浸提到葡萄酒中,而且品种对浸提率,尤其是葡萄皮缩合单宁的浸提率有更为显著的影响,这一结果与文献[20-22]研究结果一致。此外,葡萄原料中缩合单宁的浸提率较低,尤其是葡萄籽缩合单宁浸提率更低,除了受品种[9]及果皮细胞结构[17]的影响之外,还可能是由于葡萄破碎后葡萄皮及葡萄籽中浸出的缩合单宁会与果皮和果肉细胞壁多糖或者酵母细胞释放的甘露蛋白、多糖等发生结合,从而降低了其在葡萄酒中的定量[22-23]。
2.3 皮渣发酵浸渍时间对葡萄酒中缩合单宁相关味觉指标的影响
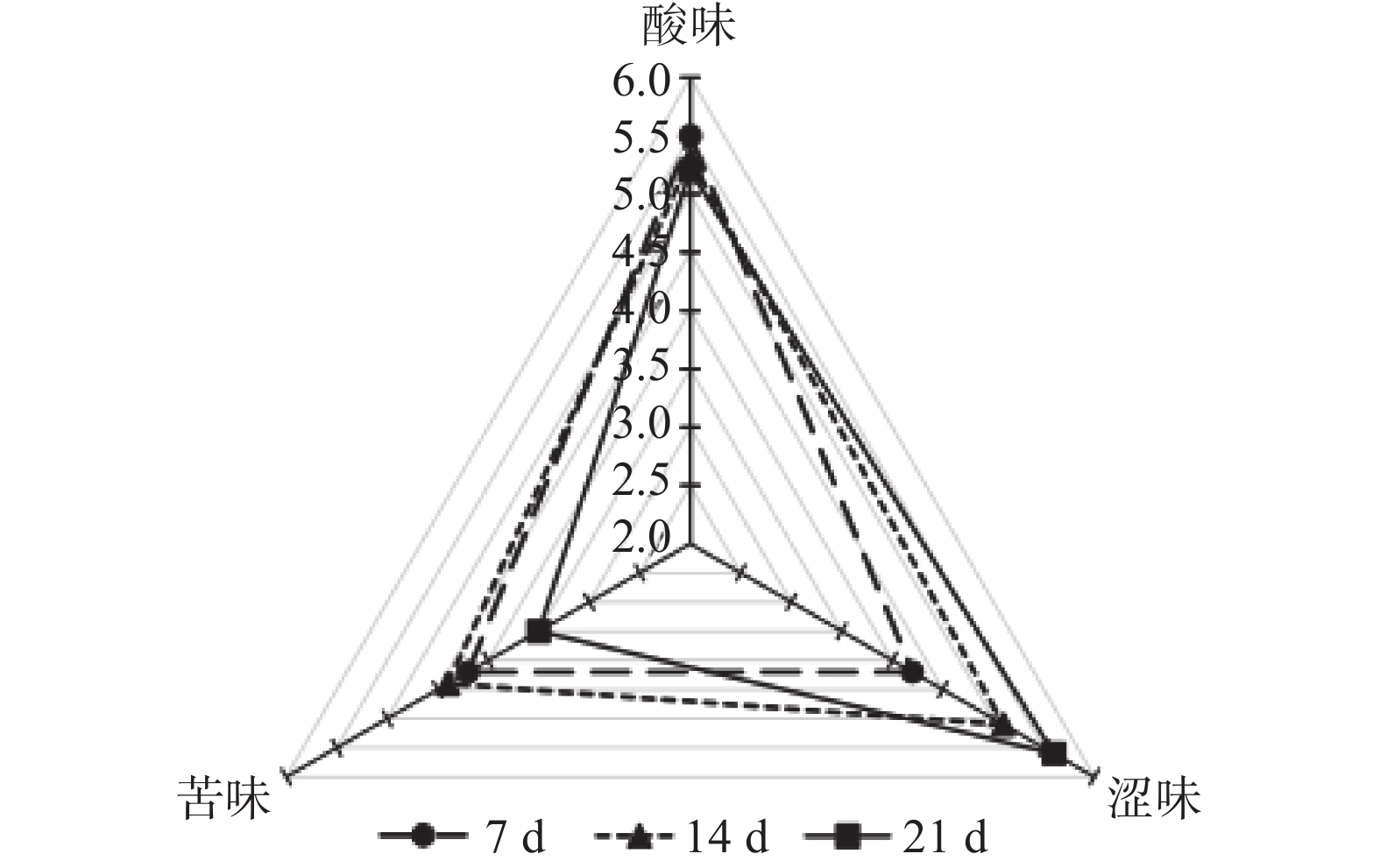
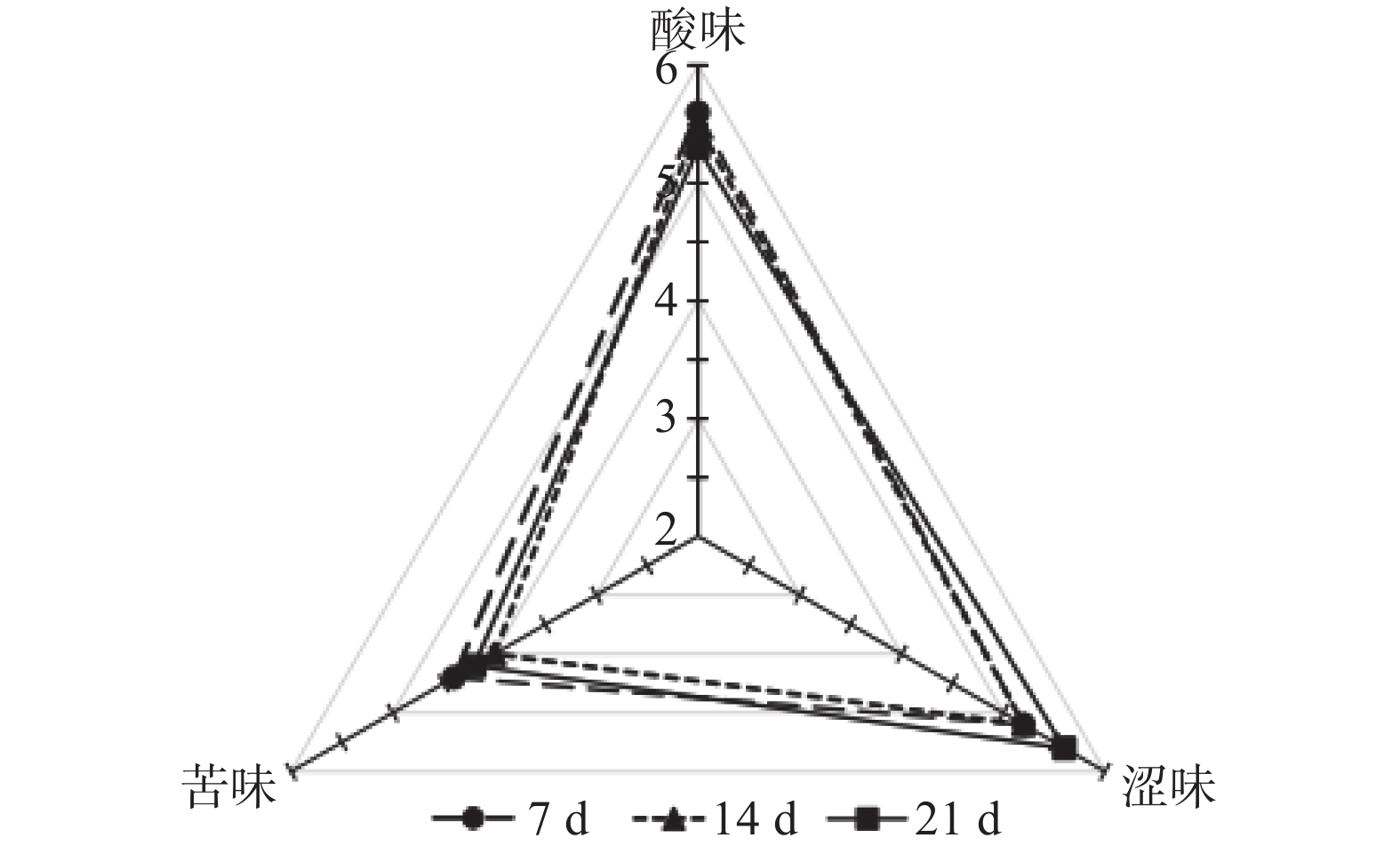
葡萄原料中酚类物质的含量及其在酒中与其他物质的互作反应与红葡萄酒的感官特性密切相关。从图1和图2中可以看出,浸渍21 d处理的葡萄酒涩味得分较高,这可能与其中缩合单宁的浓度较高有关[24-27]。不同浸渍发酵时间处理之间酸味无显著差异;浸渍21 d的赤霞珠葡萄酒的苦味与其他两个处理有显著(P<0.05)差异,西拉葡萄酒3个处理间的苦味无显著差异。由此可见,相对于苦味和酸味,浸渍时间处理对葡萄酒的涩味影响更为明显。
![]() 图 1 不同浸渍发酵时间赤霞珠葡萄酒缩合单宁相关味觉指标分析Figure 1. Mouth-feel attributes related with condensed tannins in CS wines produced with different maceration times
图 1 不同浸渍发酵时间赤霞珠葡萄酒缩合单宁相关味觉指标分析Figure 1. Mouth-feel attributes related with condensed tannins in CS wines produced with different maceration times![]() 图 2 不同浸渍发酵时间西拉葡萄酒缩合单宁相关味觉指标分析Figure 2. Mouth-feel attributes related with condensed tannins in Syrah wines produced with different maceration times
图 2 不同浸渍发酵时间西拉葡萄酒缩合单宁相关味觉指标分析Figure 2. Mouth-feel attributes related with condensed tannins in Syrah wines produced with different maceration times3. 结论
不同葡萄品种中缩合单宁的含量、组成、聚合度及没食子酰化率都有显著差异(P<0.05)。酿酒葡萄果皮中缩合单宁的平均聚合度mDP显著高于葡萄籽(P<0.05),而葡萄皮缩合单宁的没食子酰化率则显著低于葡萄籽缩合单宁(P<0.05)。葡萄皮和葡萄籽缩合单宁的组成也有所不同,其中葡萄皮缩合单宁末端单元主要是由(+)-儿茶素(CAT)组成,其次是(−)-表儿茶素(EC);而延伸单元则主要是由(−)-表棓儿茶素(EGC)组成和(−)-表儿茶素(EC)主导组成;而葡萄籽缩合单宁末端单元均主要由(+)-儿茶素(CAT)、(−)-表儿茶素(EC)组成,其次是(−)-表儿茶素没食子酸酯(ECG),而延伸单元则主要由(−)-表儿茶素(EC)组成,其次是(−)-表儿茶素没食子酸酯(ECG),不含有(−)-表棓儿茶素(EGC)延伸单元。
-
![]()
图 1 不同浸渍发酵时间赤霞珠葡萄酒缩合单宁相关味觉指标分析
Figure 1. Mouth-feel attributes related with condensed tannins in CS wines produced with different maceration times
![]()
图 2 不同浸渍发酵时间西拉葡萄酒缩合单宁相关味觉指标分析
Figure 2. Mouth-feel attributes related with condensed tannins in Syrah wines produced with different maceration times
表 1 赤霞珠和西拉葡萄皮中缩合单宁的含量及组成
Table 1 Contents and composition of condensed tannins in CS and Syrah skin
指标 赤霞珠 西拉 缩合单宁含量CTs (mg/g皮) 9.42 ± 0.31a 5.32 ± 0.12b 缩合单宁含量CTs (mg/100 g葡萄) 80.92 ± 3.10a 44.36 ± 2.48b 平均聚合度mDP 23.50 ± 2.31a 14.61 ± 1.39b 没食子酰化率G(%) 2.41 ± 0.37b 4.22 ± 0.76a 儿茶素末端单元比tCAT(%) 3.12 ± 0.05b 6.27 ± 0.09a 表儿茶素末端单元比tEC(%) 1.72 ± 0.02b 2.70 ± 0.04a 表儿茶素没食子酸酯末端单元比tECG(%) 0.14 ± 0.01a 0.16 ± 0.01a 儿茶素延伸单元比ext CAT(%) 1.78 ± 0.03a 1.71 ± 0.03a 表儿茶素延伸单元比ext EC(%) 36.61 ± 0.16b 54.40 ± 0.25a 表儿茶素没食子酸酯延伸单元比ext ECG(%) 2.01 ± 0.06b 4.27 ± 0.08a 表棓儿茶素延伸单元比ext EGC(%) 54.61 ± 0.31a 30.50 ± 0.12b 注:同行不同小写字母表示差异显著(P<0.05);表2~表3同。  下载: 导出CSV
下载: 导出CSV
表 2 赤霞珠和西拉葡萄籽中缩合单宁的含量及组成
Table 2 Content and composition of condensed tannins in CS and Syrah seed
指标 赤霞珠 西拉 缩合单宁含量CTs (mg/g籽) 24.56 ± 1.41a 20.89 ± 1.38b 缩合单宁含量CTs (mg/100 g葡萄) 99.81 ± 3.54a 87.78 ± 3.38b 平均聚合度mDP 6.41 ± 0.32a 6.22 ± 0.31a 没食子酰化率G(%) 15.82 ± 0.45b 20.65 ± 0.51a 儿茶素末端单元比tCAT(%) 7.12 ± 0.08a 6.94 ± 0.11a 表儿茶素末端单元比tEC(%) 5.20 ± 0.07b 6.28 ± 0.08a 表儿茶素没食子酸酯末端单元比tECG(%) 2.11 ± 0.05b 3.57 ± 0.06a 儿茶素延伸单元比ext CAT(%) 3.89 ± 0.07b 5.93 ± 0.10a 表儿茶素延伸单元比ext EC(%) 70.48 ± 0.65a 62.67 ± 0.56b 表儿茶素没食子酸酯延伸单元比ext ECG(%) 11.20 ± 0.15b 14.61 ± 0.17a 表棓儿茶素延伸单元比ext EGC(%) − −
下载: 导出CSV
表 3 不同浸渍时间处理赤霞珠和西拉葡萄酒中缩合单宁组成及含量
Table 3 Content and composition of condensed tannins in CS and Syrah wines produced with different maceration times
指标 赤霞珠 西拉 7 d 14 d 21 d 7 d 14 d 21 d 缩合单宁含量CTs (mg/L) 164.36 ± 10.89c 369.74 ± 19.23b 408.05 ± 22.45a 340.98 ± 18.98c 513.90 ± 28.86b 565.14 ± 29.98a 平均聚合度mDP 8.43 ± 0.22a 7.47 ± 0.20b 6.87 ± 0.18c 7.84 ± 0.21a 7.13 ± 0.18b 6.24 ± 0.16c 没食子酰化率G(%) 1.79 ± 0.12b 1.91 ± 0.12b 2.65 ± 0.15a 3.03 ± 0.18b 3.69 ± 0.16a 3.82 ± 0.18a 儿茶素末端单元比tCAT(%) 12.40 ± 0.22a 11.01 ± 0.20b 12.89 ± 0.25a 10.14 ± 0.18c 11.12 ± 0.20b 12.98 ± 0.23a 表儿茶素末端单元比tEC(%) 2.55 ± 0.03b 1.86 ± 0.01c 3.25 ± 0.02a 3.32 ± 0.03b 3.09 ± 0.02b 4.32 ± 0.04a 表儿茶素没食子酸酯末端单元比tECG(%) 0.18 ± 0.01a 0.16 ± 0.01b 0.15 ± 0.01b 0.31 ± 0.02a 0.34 ± 0.02a 0.26 ± 0.01b 儿茶素延伸单元比ext CAT(%) 6.98 ± 0.12a 4.07 ± 0.10b 3.24 ± 0.10b 1.05 ± 0.06b 1.32 ± 0.06b 1.81 ± 0.08a 表儿茶素延伸单元比ext EC(%) 39.89 ± 0.16c 42.38 ± 0.20b 44.32 ± 0.21a 50.46 ± 0.25a 50.30 ± 0.23a 49.67 ± 0.20b 表棓儿茶素延伸单元比ext EGC(%) 36.97 ± 0.19b 39.26 ± 0.24a 34.45 ± 0.22b 32.49 ± 0.20a 31.31 ± 0.22a 28.15 ± 0.16b 表儿茶素没食子酸酯延伸单元比ext ECG(%) 1.02 ± 0.01b 1.26 ± 0.02b 1.69 ± 0.02a 2.23 ± 0.06c 2.52 ± 0.08b 2.78 ± 0.07a 葡萄皮衍生缩合单宁含量(mg/L) 126.98 ± 6.56b 267.66 ± 11.22a 283.84 ± 12.23a 283.91 ± 12.89b 419.03 ± 14.56a 431.31 ± 16.87a 酒中葡萄皮衍生缩合单宁占比(%) 77.26 ± 0.32a 72.39 ± 0.28b 69.56 ± 0.20c 83.26 ± 0.27a 81.54 ± 0.26a 76.32 ± 0.22b 葡萄籽衍生缩合单宁含量(mg/L) 59.90 ± 2.56c 89.06 ± 3.10b 147.41 ± 5.23a 43.21 ± 1.89c 82.38 ± 2.89b 112.10 ± 4.01a 酒中葡萄籽衍生缩合单宁占比(%) 22.74 ± 0.18c 27.61 ± 0.21b 30.44 ± 0.22a 16.74 ± 0.12b 18.46 ± 0.14b 23.68 ± 0.20a 葡萄皮缩合单宁浸提率(%) 10.19 ± 0.09b 21.50 ± 0.12a 22.78 ± 0.12a 41.60 ± 0.29b 61.37 ± 0.28a 63.19 ± 0.34a 葡萄籽缩合单宁浸提率(%) 3.90 ± 0.10c 5.79 ± 0.13b 9.58 ± 0.16a 3.19 ± 0.10c 6.11 ± 0.12b 8.32 ± 0.17a
下载: 导出CSV
-
[1] Souquet J M, Cheynier V, Brossaud F, et al. Polymeric proanthocyanidins from grape skins[J]. Phytochemistry,1996,43(2):509−512. doi: 10.1016/0031-9422(96)00301-9
[2] Prieur C, Rigaud J, Cheynier V, et al. Oligomeric and polymeric procyanidins from grape seeds[J]. Phytochemistry,1994,36(3):781−784. doi: 10.1016/S0031-9422(00)89817-9
[3] Aron P M, Kennedy J A. Compositional investigation of phenolic polymers isolated from Vitis vinifera L. cv. Pinot Noir during fermentation[J]. Journal of Agricultural and Food Chemistry,2007,55(14):5670−5680. doi: 10.1021/jf0704817
[4] 曹鹏, 段长青. 葡萄果实和葡萄酒中缩合单宁的研究进展[J]. 农业工程学报,2004,20(增刊):5−12. [5] 张振文, 宁鹏飞, 张军贤, 等. 葡萄酒缩合单宁测定方法的比较研究[J]. 食品科学,2012,33(20):233−237. [6] Kennedy J A, Hayasaka Y, Vidal S, et al. Composition of grape skin proanthocyanidins at different stages of berry development[J]. Journal of Agricultural and Food Chemistry,2001,49(11):5348−5355. doi: 10.1021/jf010758h
[7] Peyrot des Gachons C, Kennedy J A. Direct method for determining seed and skin proanthocyanidin extraction into red wine[J]. Journal of Agricultural and Food Chemistry,2003,51(20):5877−5881. doi: 10.1021/jf034178r
[8] Harbertson J F, Kennedy J A, Adams D O. Tannin in skins and seeds of Cabernet Sauvignon, Syrah, and Pinot noir berries during ripening[J]. American Journal of Enology and Viticulture,2002,53(1):54−59.
[9] Romero-Cascales I, Ortega-Regules A, López-Roca J M, et al. Differences in anthocyanin extractability from grapes to wines according to variety[J]. American Journal of Enology and Viticulture,2005,56(3):212−219.
[10] Busse-Valverde N, Gómez-Plaza E, López-Roca J M, et al. Effect of different enological practices on skin and seed proanthocyanidins in three varietal wines[J]. Journal of Agricultural and Food Chemistry,2010,58(21):11333−11339. doi: 10.1021/jf102265c
[11] López N, Puértolas E, Hernández-Orte P, et al. Effect of a pulsed electric field treatment on the anthocyanins composition and other quality parameters of Cabernet Sauvignon freshly fermented model wines obtained after different maceration times[J]. LWT-Food Science and Technology,2009,42(7):1225−1231. doi: 10.1016/j.lwt.2009.03.009
[12] 李华, 王华, 袁春龙, 等. 葡萄酒工艺学[M]. 北京: 科学出版社, 2007. [13] Kennedy J A, JonesGP. Analysis of proanthocyanidin cleavage products following acid-catalysis in the presence of excess phloroglucinol[J]. Journal of Agricultural and Food Chemistry,2001,49(4):1740−1746. doi: 10.1021/jf001030o
[14] Pastor del Rio J L, KennedyJA. Development of proanthocyanidins in Vitis vinifera L. cv. Pinot noir grapes and extraction into wine[J]. American Journal of Enology and Viticulture,2006,57(2):125−132.
[15] Ducasse M A, Canal-Llauberes R M, de Lumley M, et al. Effect of macerating enzyme treatment on the polyphenol and polysaccharide composition of red wines[J]. Food Chemistry,2010,118(2):369−376. doi: 10.1016/j.foodchem.2009.04.130
[16] Cosme F, Ricardo-da-Silva J M, Laureano O, et al. Tannin profiles of Vitis vinifera L. cv. red grapes growing in Lisbon and from their monovarietal wines[J]. Food Chemistry,2009,112(1):197−204. doi: 10.1016/j.foodchem.2008.05.058
[17] Ortega-Regules A, Romero-Cascales I, Ros-García J M, et al. A first approach towards the relationship between grape skin cell-wall composition and anthocyanin extractability[J]. Analytica Chimica Acta,2006(1-2):26−32.
[18] Hernández-Hierro J M, Quijada-Morín N, Martínez-Lapuente L, et al. Relationship between skin cell wall composition and anthocyanin extractability of Vitis vinifera L. cv. Tempranillo at different grape ripeness degree[J]. Food Chemistry,2014,146:41−47. doi: 10.1016/j.foodchem.2013.09.037
[19] Quijada-Morín N, Hernández-Hierro J M, Rivas-Gonzalo J C, et al. Extractability of low molecular mass flavanols and flavonols from red grape skins. Relationship to cell wall composition at different ripeness stages[J]. Journal of Agricultural and Food Chemistry,2015,63(35):7654−7662. doi: 10.1021/acs.jafc.5b00261
[20] González-Manzano S, Rivas-Gonzalo J C, Santos-Buelga C, et al. Extraction of flavan-3-ols from grape seed and skin into wine using simulated maceration[J]. Analytica Chimica Acta,2004,513(1):283−289. doi: 10.1016/j.aca.2003.10.019
[21] Douglas O, Adams. Phenolics and ripening in grape berries[J]. American Journal of Enology and Viticulture,2006,57(3):249−256.
[22] Bindon K A, Smith P A, Holt H, et al. Interaction between Grape-derived proanthocyanidins and cell wall material. 2. Implications for vinification[J]. Journal of Agricultural and Food Chemistry,2010,58(19):10736−10746. doi: 10.1021/jf1022274
[23] Vidal S, Francis L, Williams P, et al. The mouth-feel properties of polysaccharides and anthocyanins in a wine like medium[J]. Food Chemistry,2004,85:519−525. doi: 10.1016/S0308-8146(03)00084-0
[24] Obreque-Slier E, Peña-Neira Á, López-Solís R, et al. Interactions of enological tannins with the protein fraction of saliva and astringency perception are affected by pH[J]. LWT-Food Science and Technology,2012,45(1):88−93. doi: 10.1016/j.lwt.2011.07.028
[25] Villamor R R, Harbertson J F, Ross C F, et al. Influence of tannin concentration, storage temperature, and time on chemical and sensory properties of Cabernet Sauvignon and Merlot wines[J]. American Journal of Enology and Viticulture,2009,60(4):442−449.
[26] Fontoin H, Saucier C, Teissedre P L, et al. Effect of pH, ethanol and acidity on astringency and bitterness of grape seed tannin oligomers in model wine solution[J]. Food Quality and Preference,2008,19(3):286−291. doi: 10.1016/j.foodqual.2007.08.004
[27] Singleton V L, Sieberhagen H A, De Wet P, et al. Composition and sensory qualities of wines prepared from white grapes by fermentation with and without grape solids[J]. American Journal of Enology and Viticulture,1975,26(2):62−69.
 下载:
下载:


计量
- 文章访问数:
- HTML全文浏览量:
- PDF下载量:
