


Protective Effects of Giant Salamander Bioactive Peptides on D-galactose Induced Oxidative Damage in Mice
-
摘要: 为了探究大鲵活性肽抗衰老作用机制,利用非标记定量蛋白质组方法对去除高丰度蛋白质后的灌胃大鲵活性肽小鼠血清进行蛋白质组生物信息学分析。结果表明,去除高丰度蛋白质后的血清中共检测到425种蛋白质,其中26种蛋白质表达量变化显著(P<0.05)。这些差异表达的低丰度蛋白质参与14个生理过程、具有6种分子功能、存在于7种细胞组分中。其中胞质四氢叶酸合成酶和肌糖原磷酸化酶显著增加(P<0.05),分别增加了1.78和1.05倍,黄嘌呤脱氢酶显著减少(P<0.05),这些酶表达量的增加或降低,可以减少体内自由基的量。综上,大鲵活性肽抗衰老的机制是减少机体内自由基而发挥作用,本研究为大鲵活性肽的开发利用提供了基础数据。Abstract: To investigate anti-aging mechanism of giant salamander bioactive peptides, label-free quantitative (LFQ) proteomics was used after high-abundant proteins were removed. The results of LFQ showed 425 low-abundant proteins in serum in mice, in which 26 proteins were differentially expressed (P<0.05), and covered 14 biological processes, 6 molecular functions and 7 cellular components. Tetrahydrofolate synthase and glycogen phosphorylase were up-regulated expressed significantly (P<0.05), increased by 1.78 times and 1.05 times respectively, while xanthine dehydrogenase was down-regulated expressed significantly (P<0.05) decreased. The expression of these enzymes increased or decreased, which could reduce the amount of free radicals in vivo. In summary, the giant salamander bioactive peptides prevented aging by scavenging free radicals, this study would provide basic data for the use of the giant salamander bioactive peptides.
-
大鲵(Andrias davidianus),属于两栖纲、有尾目、隐鳃鲵科,是我国特有的珍稀两栖动物,属于国家二类保护动物[1]。大鲵不仅有丰富的营养,而且还有重要的药用价值。据《本草纲目》记载[2]:“鲵鱼,甘,有毒。食之无痴疾。”1978年出版的《全国中草药汇编》将大鲵列为药品,主要治疗神经衰弱、病后体虚、贫血、痢疾等疾病[3]。研究表明,大鲵含有丰富的蛋白质、不饱和脂肪酸、多糖、矿物质元素等成分[4]。这些营养物质构成了大鲵活性成分的物质基础。大鲵肌肉中蛋白质含量高,氨基酸比例合理,种类丰富。大鲵肌肉蛋白质经过酶解过程可以获得具有抗氧化、免疫调节等生物活性的大鲵活性肽[5-7],适用于大鲵肽相关产品的开发。
衰老是多细胞生物体在发育成熟后受多因素作用的生理机能退行性过程,主要有自由基、免疫、端粒细胞与神经内分泌等学说解释机体的衰老原因,但主流的解释为自由基攻击等氧化反应导致机体衰老[8]。因机体内脏器受氧化物攻击,产生有毒有害物质无法及时清除,产生一系列生理生化应激反应,进而导致表观上、生理上的退行性衰老。研究表明,长期大量对动物注射D-半乳糖溶液,过多的D-半乳糖在醛糖还原酶的作用下生成半乳糖醇,半乳糖醇不能被机体代谢,堆积在细胞内的半乳糖醇可以导致细胞的氧化应激反应,进而导致细胞内活性氧(reactive oxygen species, ROS)增加[9]。过多的ROS引起细胞损害,导致机体多器官、多系统功能减退[10-11]。D-半乳糖可以使机体的很多组织出现类似衰老的退行性改变,出现与衰老相关的毛色变暗、行动迟缓等体征、生化改变、免疫功能下降、退行性神经功能紊乱,还出现胰腺损伤、脑损伤以及肾脏损伤[2-15]。因此,D-半乳糖致小鼠衰老模型被广泛用来评价各种生物活性成分的抗衰老作用。
目前,对大鲵肌肉的营养成分进行分析的研究报道较多。对大鲵肽的酶解制备工艺及其具有抗氧化活性的研究也有报道[16-17]。但鲜有对大鲵肽的抗衰老作用的深入研究。本实验采用D-半乳糖皮下注射建立小鼠亚急性衰老模型以及非定量标记蛋白质组学技术研究大鲵肽的抗衰老作用,以期从分子水平上阐明大鲵活性肽抗衰老机理,为大鲵的高值化开发利用提供科学依据。
1. 材料与方法
1.1 材料与仪器
雄性SPF小鼠 体重18~22 g,大连医科大学实验动物中心,生产许可证SCXK(辽)2013-0003;大鲵活性肽 分子量1400~2000 Da,由辽宁省水产品加工及综合利用重点实验室制备;D-半乳糖 国药集团化学试剂有限公司;总抗氧化能力(Total antioxidant capacity,T-AOC)、超氧化物歧化酶(Superoxide dismutase,SOD)、丙二醛(Malondialdehyde,MDA)测定试剂盒 南京建成生物工程研究所。
5430R低温高速离心机、真空离心浓缩仪 Eppendorf公司;Q exactive质谱仪 Thermo Scientific公司;Triple TOF 6600+质谱仪 AB SCIEX公司;Agilent 1290 Infinity LC超高压液相色谱仪 Agilent公司。
1.2 实验方法
1.2.1 D-半乳糖致小鼠衰老模型的建立、给药和处理
将60只SPF级昆明小鼠(雄性)分为5组,分别为正常组、衰老模型组,大鲵活性肽低、中、高剂量组,对应剂量分别为:100、200、400 mg/(kg·d)。除正常组每天注射等体积生理盐水外,其他各组每天都单次皮下组织注射5% D-半乳糖0.5 mL/d(生理盐水配制)。每周记录1次小鼠体重。大鲵活性肽低、中、高剂量组每日按0.1 mL大鲵活性肽水溶液/20 g小鼠体重分别灌胃一次,正常组和衰老模型组以生理盐水代替大鲵活性肽水溶液进行灌胃,同时观察小鼠毛色与行动。连续灌胃6周后,称重,摘取眼球取血,立即离心(3000 r/min,10 min),取上层血清,−20 ℃冰箱冻存备用。
1.2.2 小鼠组织脏器指数测定
小鼠称重后处死,快速取出脑、肝脏、胸腺和脾4种脏器,用滤纸吸干血水,电子天平上称重。按照式(1)计算肝脏指数、脑指数、胸腺指数和脾指数。
脏器指数(mg/10g)=脏器质量(mg)/小鼠体质量(10g) (1) 1.2.3 生化指标测定
小鼠血清中总抗氧化(T-AOC)能力、超氧化物歧化酶(SOD)活性和丙二醛(MDA)含量的测定,按试剂盒说明书的操作步骤进行。
1.2.4 蛋白质组样品制备
取50 μL大鲵活性肽中剂量组和D-半乳糖致小鼠衰老模型组小鼠血清,均采用去血清高丰度亲和色谱柱Agilent Multiple Affinity Removal LC Column-Mouse 3(4.6 mm×100 mm,1.66 mL),获得小鼠血清中低丰度蛋白组分。各样品血清低丰度蛋白组分经过超滤浓缩后加入等体积SDT裂解液,沸水浴裂解15 min,于14000×g条件下离心20 min。采用BCA法对上清液进行定量。大鲵活性肽中剂量组小鼠血清样品标记为ADB-T,D-半乳糖致小鼠衰老模型组小鼠血清样品标记为ADB-M。
1.2.5 SDS-PAGE电泳
取裂解后的各样品分别取20 μg,分别加入等体积2×上样缓冲液,沸水浴5 min,取20 μL进行12.5% SDS-PAGE电泳后,考马斯亮蓝染色后脱色液脱色,判断样品裂解情况。
1.2.6 样品酶解
采用Filter aided proteome preparation (FASP)方法对各样品进行酶解[18]。分别取样品ADB-T及ADB-M各30 μL蛋白质溶液,分别加入0.003 mmol DTT,沸水浴5 min后冷却至室温。加入200 μL尿素缓冲液(8 mol/L尿素,150 mmol/L Tris-HCl,pH8.0)混匀后,在10 kD超滤离心管中于14000×g离心15 min,弃滤液(重复该步骤一次)。加入100 μL 100 mmol/L IAA缓冲液(以尿素缓冲液溶解),在600 r/min条件下振荡1 min,室温避光30 min后于相同离心条件下进行离心。加入100 μL尿素缓冲液,相同离心条件下离心,该步骤重复两次。加入100 μL 25 mmol/L NH4HCO3溶液,相同离心条件下离心两次。加入40 μL 0.1 g/L胰蛋白酶缓冲液(以100 mmol/L NH4HCO3溶液配制),600 r/min振荡反应1 min,37 ℃静置16 h。换新收集管,相同离心条件下离心;再加入40 μL 25 mmol/L NH4HCO3,在相同离心条件下离心并收集滤液。通过C18 Cartridge脱盐冻干后以40 μL 0.1%甲酸溶液复溶,测定280 nm吸光值[19]。
1.2.7 LC-MS/MS分析
根据FASP酶解后OD280定量结果,各取2 μg酶解后的ADB-T及ADB-M进行LC-MS/MS分析。样品先经过RP-C18毛细管高效液相色谱柱(Thermo EASY column SC200 150 μm×100 mm)的纳升流速HPLC液相系统EASY-nLC1000,以甲酸乙腈水溶液进行梯度分离。预柱为RP-C18 Thermo EASY column SC001 traps 150 μm×20 mm。A液为0.1%甲酸乙腈水溶液(乙腈为2%),B液为0.1%甲酸乙腈水溶液(乙腈为84%)。梯度条件为:0~100 min,B液线性梯度0%~45%;100~108 min,B液线性梯度45%~100%;108~120 min,B液维持在100%[19]。分离后用Q-Exactive质谱仪进行质谱分析[15]。预分析时长60 min,正式分析时长120 min。
1.2.8 MaxQuant的数据分析
质谱分析原始数据为RAW文件,采用MaxQuant软件(版本号1.3.0.5)[20-21]进行查库鉴定,并根据Label free算法进行定量分析[22]。
1.2.9 Gene Ontology(GO)功能注释
利用Blast2GO[23]对目标蛋白质集合进行GO注释。利用NCBI BLAST+ (ncbi-blast-2.2.28+-win32.exe)比对小鼠血清数据库,保留E值小于等于0.001的前10条。再依次通过提取、注释。
1.2.10 KEGG通路注释
KEGG Automatic Annotation Server软件[24]获取目标蛋白质序列参与的通路信息。通过Fisher精确检验评价GO term或KEGG通路蛋白质富集度的显著性水平。
1.2.11 蛋白质聚类分析(Clustering)
对目标蛋白质定量信息归一化处理后,进行蛋白质聚类分析。
1.3 数据处理
数据处理采用SPSS 19.0软件进行单因素方差分析,结果表示为平均值±标准差,P<0.05为差异显著。
2. 结果与分析
2.1 小鼠体质变化情况及脏器指数
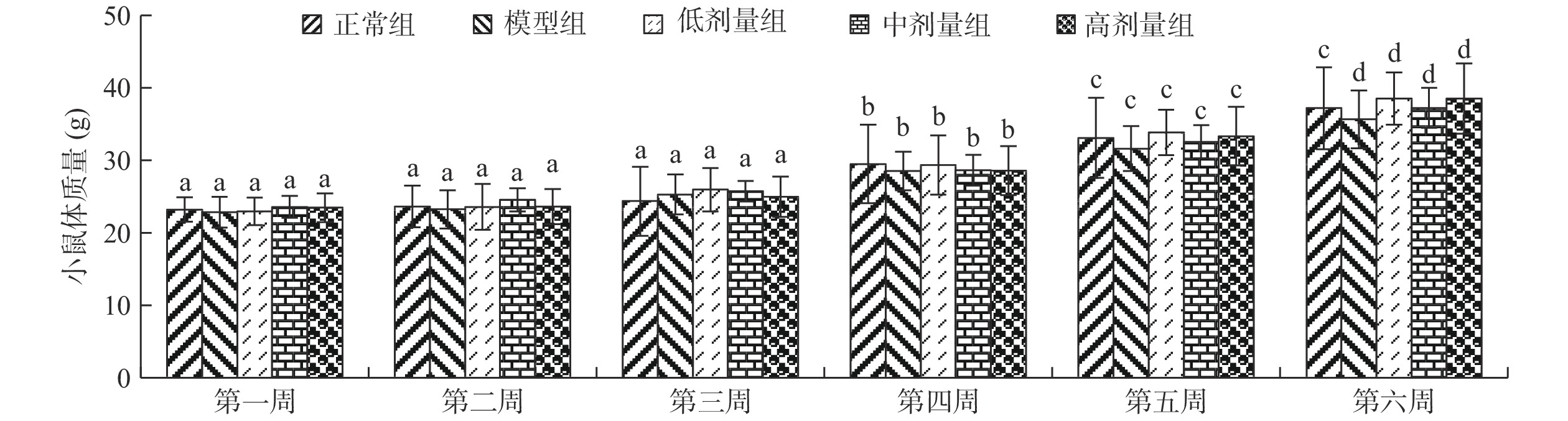
各组小鼠在实验前,皮毛光滑,喜攀爬、啮咬树枝。衰老模型组小鼠连续6周每天皮下组织注射5% D-半乳糖0.5 mL后,小鼠毛色变暗,行动迟缓,攀爬行为减少等变化。该结果与黄杰等[13]建立D-半乳糖致衰老小鼠的反应与状态变化结果相似。衰老模型组小鼠体重仅增加56.18%,正常小鼠体重增60.30%,增加量低于正常组(图1)。灌胃低、中、高浓度大鲵活性肽水溶液各组小鼠平均体重高于模型组,趋近于正常组,但未表现出剂量依赖性,表明大鲵活性肽在低剂量情况下具有促进衰老小鼠恢复生长作用。
![]() 图 1 大鲵活性肽对小鼠体质量的影响注:同组不同小写字母,表示为同组不同时间差异显著(P<0.05);同组相同小写字母,表示为同组不同时间差异不显著(P>0.05)。Figure 1. Effect of giant salamander bioactive peptides on body weight of mice
图 1 大鲵活性肽对小鼠体质量的影响注:同组不同小写字母,表示为同组不同时间差异显著(P<0.05);同组相同小写字母,表示为同组不同时间差异不显著(P>0.05)。Figure 1. Effect of giant salamander bioactive peptides on body weight of mice各组SPF级雄性昆明小鼠灌胃对应大鲵活性肽溶液6周后,小鼠脏器指数变化如表1。由表1可以看出,各灌胃大鲵活性肽组的脑指数、胸腺指数与正常组相比不显著(P>0.05);中剂量组的脾指数显著高于正常组(P<0.05);低、高剂量大鲵活性肽组的肝指数均极显著低于模型组(P<0.01)。灌胃大鲵肽后,小鼠的脾脏指数增加,表明大鲵肽在一定程度上可以增强小鼠机体免疫功能。
表 1 大鲵活性肽对小鼠脏器指数的影响Table 1. Effect of giant salamander bioactive peptides on organ indexes for each group of mice分组 脑指数
(mg/10 g)肝脏指数
(mg/10 g)胸腺指数
(mg/10 g)脾指数
(mg/10 g)正常组 98.8±26.4 393.5±39.9 20.5±12.2 25.2±8.3 模型组 99.4±24.0 462.3±15.8* 21.9±9.6 31.9±8.8 低剂量组 91.1±16.0 399.9±55.2## 24.4±7.5 30.9±4.5 中剂量组 94.1±15.5 454.4±25.6* 22.2±4.3 32.9±8.0* 高剂量组 87.9±17.2 415.2±30.6## 23.4±14.0 28.6±6.5 注:*表示与正常组相比,差异显著(P<0.05);**表示与正常组相比,差异极显著(P<0.01);#表示与模型组相比,差异显著(P<0.05);##表示与模型组相比,差异极显著(P<0.01);表2同。 2.2 生化指标检测
对小鼠血清中MDA含量、SOD活力和T-AOC进行测定,结果见表2。由表2可见,与模型组相比,灌胃大鲵活性肽中、高剂量组小鼠血清中MDA含量极显著降低(P<0.01),并且SOD活力和T-AOC显著升高(P<0.05)。注射D-半乳糖致小鼠衰老后,小鼠血清中MDA含量极显著上升,SOD活力和T-AOC极显著降低(P<0.01),而在灌胃大鲵活性肽后,显著改善小鼠这些生化指标,但未发现剂量依赖性。因此,大鲵活性肽具有抗D-半乳糖致小鼠衰老作用。
表 2 大鲵活性肽对小鼠生化指标的影响Table 2. Effect of giant salamander bioactive peptides on the levels of serum markers in SPF mice分组 MDA(nmol/mL) SOD活力(U/mL) T-AOC(mmol/L) 正常组 10.58±1.88 157.26±16.15 0.58±0.04 模型组 16.70±2.60** 119.88±17.49** 0.48±0.05** 低剂量组 14.86±2.10** 131.35±13.99** 0.50±0.05** 中剂量组 11.90±2.60## 141.16±16.93*## 0.54±0.05*# 高剂量组 12.08±2.27## 138.51±15.59*# 0.54±0.04*# 2.3 血清样品SDS-PAGE和质谱检测
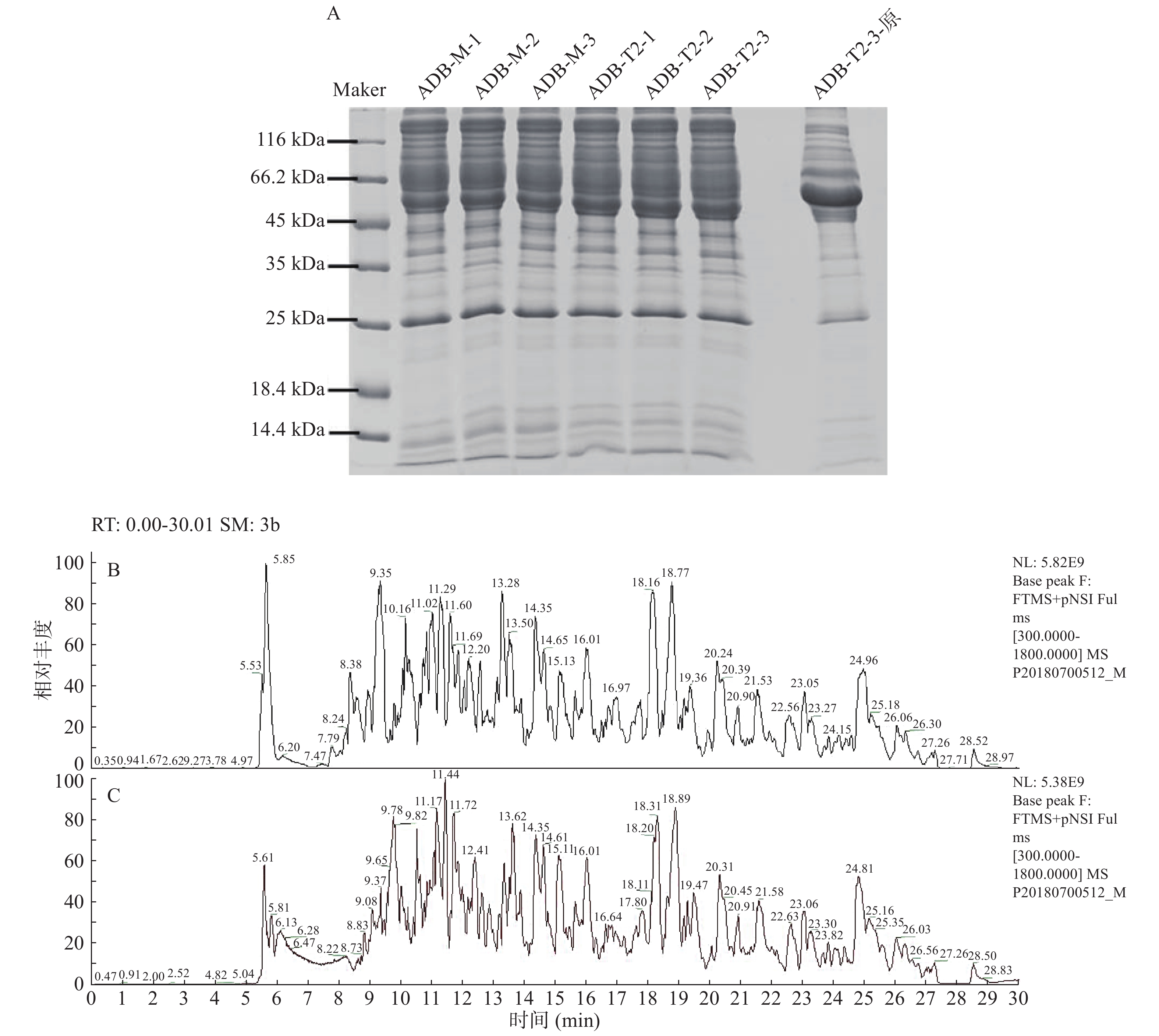
SDS-PAGE(图2A)结果表明,未经去除血清高丰度亲和色谱柱处理的中剂量组小鼠血清蛋白(ADB-T2-3原)中60 kDa附近蛋白含量过高而影响低丰度蛋白质分析,而去除高丰度血清蛋白后模型组(ADB-M-1、ADB-M-2、ADB-M-3)和的大鲵活性肽中剂量组(ADB-T2-1、ADB-T2-2、ADB-T2-3)电泳条带清晰,平行性好,可进行质谱分析。质谱分析Basepeak结果(图2B和图2C)表明,组间样本平行性好,酶解效果良好,可进行非标记定量质谱测定。
![]() 图 2 小鼠血清样本平行性注:A:小鼠血清蛋白SDS-PAGE,其中Marker为标准蛋白,ADB-M-1、ADB-M-2、ADB-M-3为模型组去除高丰度蛋白血清三个平行样,ADB-T2-1、ADB-T2-2、ADB-T2-3为中剂量组去除高丰度蛋白血清三个平行样,ADB-T2-3-原为中剂量组血清;B:去除高丰度蛋白的模型组小鼠血清蛋白质谱图;C:去除高丰度蛋白的中剂量组小鼠血清蛋白质谱图。Figure 2. Samples parallelism of proteins in serum in mice
图 2 小鼠血清样本平行性注:A:小鼠血清蛋白SDS-PAGE,其中Marker为标准蛋白,ADB-M-1、ADB-M-2、ADB-M-3为模型组去除高丰度蛋白血清三个平行样,ADB-T2-1、ADB-T2-2、ADB-T2-3为中剂量组去除高丰度蛋白血清三个平行样,ADB-T2-3-原为中剂量组血清;B:去除高丰度蛋白的模型组小鼠血清蛋白质谱图;C:去除高丰度蛋白的中剂量组小鼠血清蛋白质谱图。Figure 2. Samples parallelism of proteins in serum in mice2.4 蛋白质鉴定结果统计
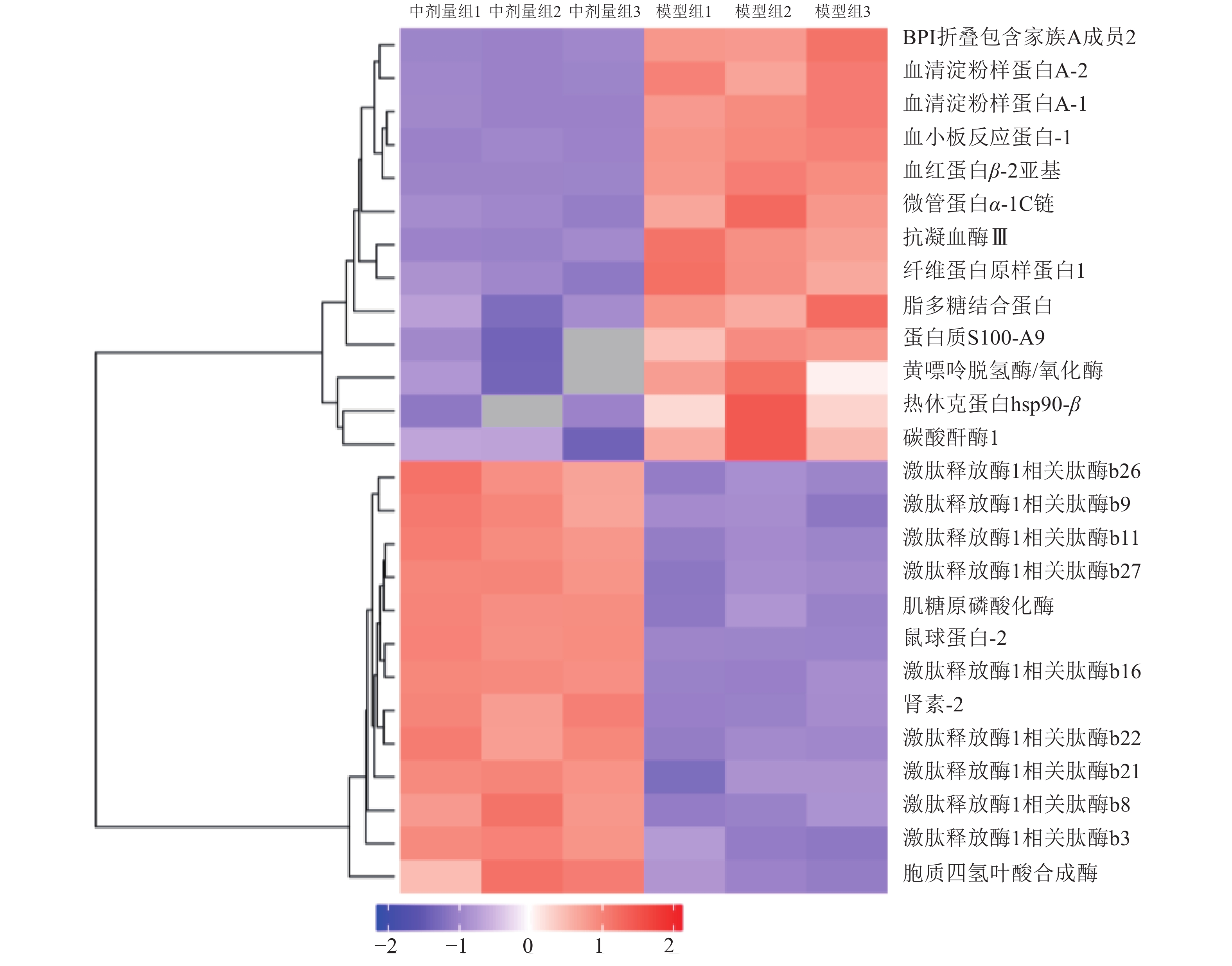
模型组与中剂量组共鉴定到蛋白质425个,其中差异表达蛋白26个,通过大鲵活性肽治疗后,差异表达蛋白质表达上调13个(表3),下调13个(表4)。其中胞质四氢叶酸合成酶(存取编号:Q922D8)和肌糖原磷酸化酶显著增加,黄嘌呤脱氢酶(氧化酶)(存取编号:Q00519)显著减少。四氢叶酸作为生物体中的一碳单位转移酶的辅酶,在氨基酸代谢和核酸代谢中起重要作用。糖原磷酸化酶催化糖原转变为1-磷酸葡萄糖,补充体液中葡萄糖。黄嘌呤脱氢酶在核酸代谢过程中起重要作用,可催化黄嘌呤转变为尿酸。
表 3 上调的差异表达蛋白质Table 3. Up-regulated proteins with significant differences存取编号 蛋白质 分子量(kDa) 中剂量组表达量/
模型组表达量t检验P值 pI 参与代谢途径 P07628 激肽释放酶1相关肽酶b8 28.531 4.49 1.43×10−4 8.28 内分泌和其它因素调节的钙再吸收
肾素-血管紧张素系统Q9JM71 激肽释放酶1相关肽酶b27 28.742 4.18 2.75×10−5 8.86 P00796 肾素-2 44.282 3.41 3.49×10−5 6.01 肾素-血管紧张素系统/肾素分泌 Q61759 激肽释放酶1相关肽酶b21 28.69 2.93 1.80×10−4 7.05 内分泌和其它因素调节的钙再吸收
肾素-血管紧张素系统Q922D8 胞质四氢叶酸合成酶 101.2 2.78 1.24×10−3 6.7 叶酸一碳库 P15946 激肽释放酶1相关肽酶b11 28.727 2.70 2.15×10−5 6.69 内分泌和其它因素调节的钙再吸收
肾素-血管紧张素系统P00756 激肽释放酶1相关肽酶b3 28.998 2.54 1.00×10−4 6.37 内分泌和其它因素调节的钙再吸收
肾素-血管紧张素系统P15948 激肽释放酶1相关肽酶b22 28.384 2.52 5.15×10−5 6.2 内分泌和其它因素调节的钙再吸收
肾素-血管紧张素系统P15949 激肽释放酶1相关肽酶b9 28.9 2.45 1.54×10−4 7.56 内分泌和其它因素调节的钙再吸收
肾素-血管紧张素系统P04071 激肽释放酶1相关肽酶b16 28.722 2.18 2.54×10−6 5.4 P36369 激肽释放酶1相关肽酶b26 28.463 2.12 2.06×10−4 6.41 内分泌和其它因素调节的钙再吸收
肾素-血管紧张素系统P28666 鼠球蛋白-2 162.38 2.12 8.12×10−7 6.29 Q9WUB3 肌糖原磷酸化酶 97.285 2.05 3.37×10−5 6.65 胰岛素信号通路/淀粉和蔗糖代谢/坏死性凋亡
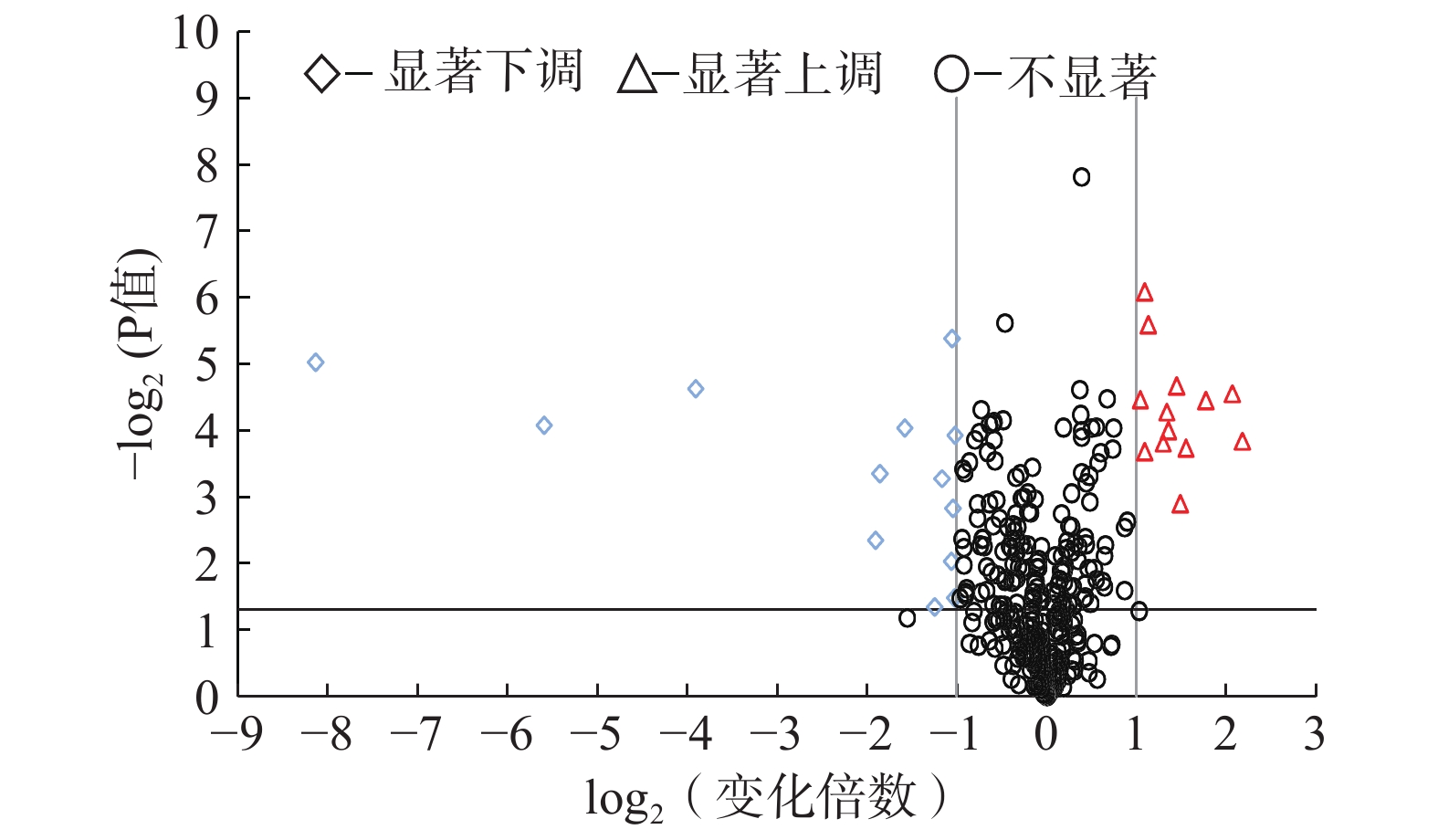
胰岛素抵抗/胰高血糖素信号通路注:中剂量组表达量/模型组表达量-表示中剂量治疗组和模型组蛋白质表达量平均值的比值;表4同。 表 4 下调的差异表达蛋白质Table 4. Down-regulated proteins with significant differences存取编号 蛋白质 分子量(kDa) 中剂量组表达量/模型组表达量 t检验P值 pI 参与代谢途径 P32261 抗凝血酶Ⅲ 52.003 0.49 1.16×10−4 6.1 补体和凝血级联反应 Q00519 黄嘌呤脱氢酶/氧化酶 146.56 0.49 3.26×10−2 7.62 过氧化物酶体/药物代谢-其他酶/嘌呤代谢/咖啡因代谢 Q61805 脂多糖结合蛋白 53.055 0.48 1.46×10−3 8.64 NF-κB信号通路/结核/Toll样受体信号通路/沙门氏菌感染 P35441 血小板反应蛋白-1 129.65 0.48 4.09×10−6 4.72 P13634 碳酸酐酶1 28.33 0.48 9.12×10−3 6.44 氮代谢 P68373 微管蛋白α-1C链 49.909 0.44 5.26×10−4 4.96 吞噬体/紧密连接/缝隙连接/凋亡 P11499 热休克蛋白hsp90-β 83.28 0.42 4.46×10−2 4.96 Th17细胞分化/坏死下垂/雌激素信号通路/内质网蛋白质处理/PI3K-Akt信号通路/抗原处理和提呈/IL-17信号通路/NOD样受体信号通路/癌症/孕酮介导的卵母细胞成熟/前列腺癌/流体剪切应力和动脉粥样硬化的途径 P07743 BPI折叠包含家族A成员2 24.753 0.33 9.06×10−5 4.87 Q71KU9 纤维蛋白原样蛋白1 36.439 0.28 4.40×10−4 5.48 P31725 蛋白质S100-A9 13.049 0.27 4.49×10−3 6.65 IL-17信号通路 P05366 血清淀粉样蛋白A-1 13.77 0.07 2.30×10−5 6.5 P05367 血清淀粉样蛋白A-2 13.622 0.02 8.21×10−5 6.4 P02089 血红蛋白β-2亚基 15.878 0.00 9.12×10−6 7.85 以治疗组对模型组表达倍数与t检验P值作图,显示两组间蛋白质表达差异显著性的火山图如图3所示,图中横坐标大于1且纵坐标大于1.3的区域为蛋白质表达显著上调区域,横坐标小于−1且纵坐标大于1.3的区域为蛋白质表达显著下调区域。从图3中可以看出,13个表达上调的蛋白质中,最大表达倍数为4倍多(log24.49),而表达下调的13个蛋白质中有3个表达下调量极大,分别为0.067、0.021、0.004倍。
对26个差异表达蛋白质进行聚类分析,结果如图4所示。这些差异表达蛋白质主要分为两类,即表达量上调和下调。在MCL-T处理组的上调表达的蛋白质中,胞质四氢叶酸合成酶在各组中的差异较大,在下调表达的蛋白质中,热休克蛋白HSP 90-β亚基、黄嘌呤脱氢酶和碳酸酐酶在各组中表达量差异较大。
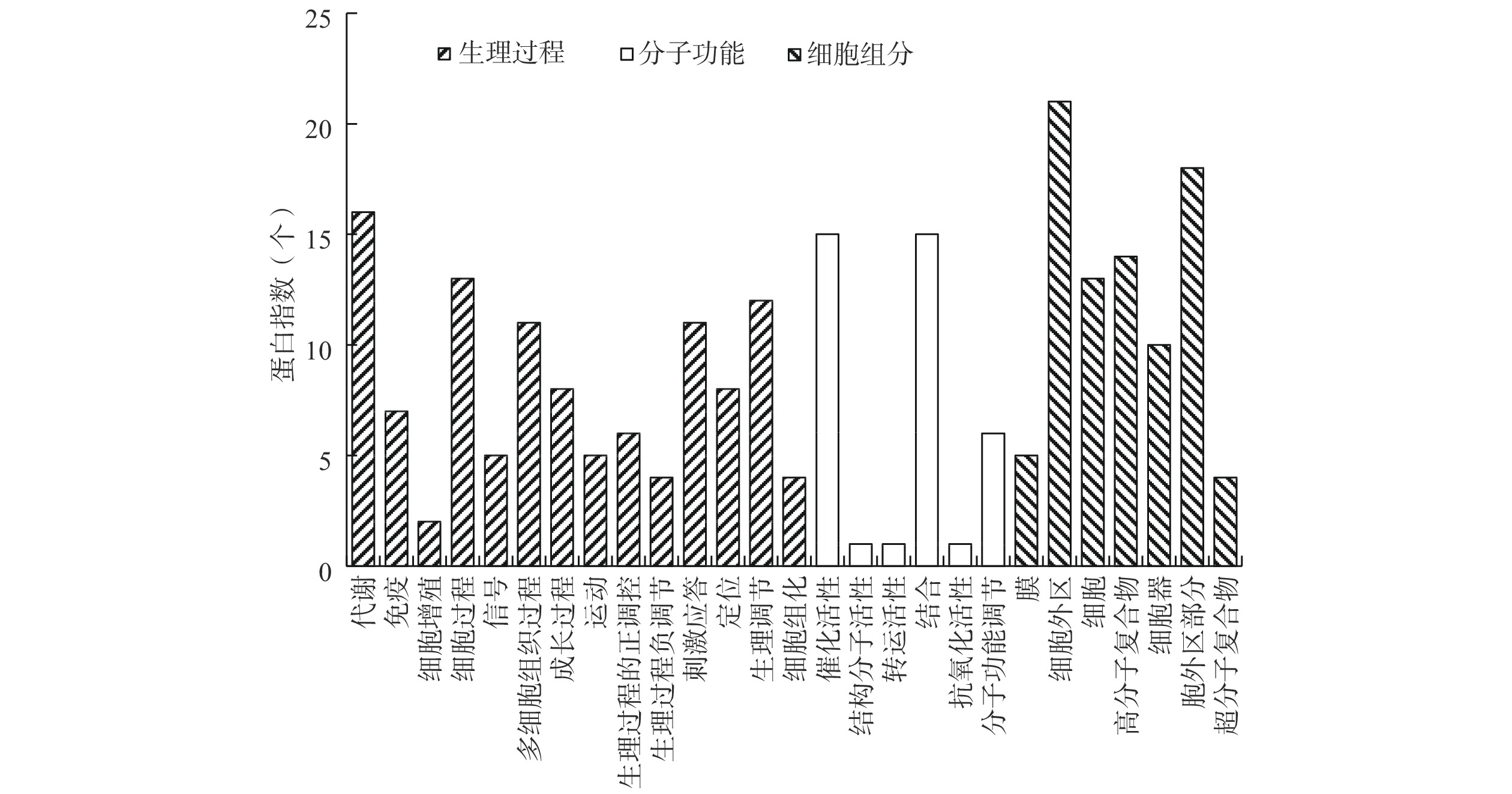
通过Blast2Go(https://www.blast2go.com/)软件对鉴定到的425种蛋白质进行基因本体(Gene ontology,GO)功能注释,2水平注释结果如图5所示。从图5中可以看出,小鼠低丰度蛋白参与了14个生理过程、具有6种分子功能、存在于7种细胞组分中。在这些生理过程中,代谢过程、细胞生成过程、多细胞组织过程、刺激应答、生理调节等过程都有10个以上差异表达蛋白;这些差异表达蛋白中,有15个具有催化活性、15个具有结合功能、6个具有分子功能调节活性;其中胞外区有21个、细胞中有13个、高分子复合物有14个、细胞器中有10个、胞外区部分有18个。
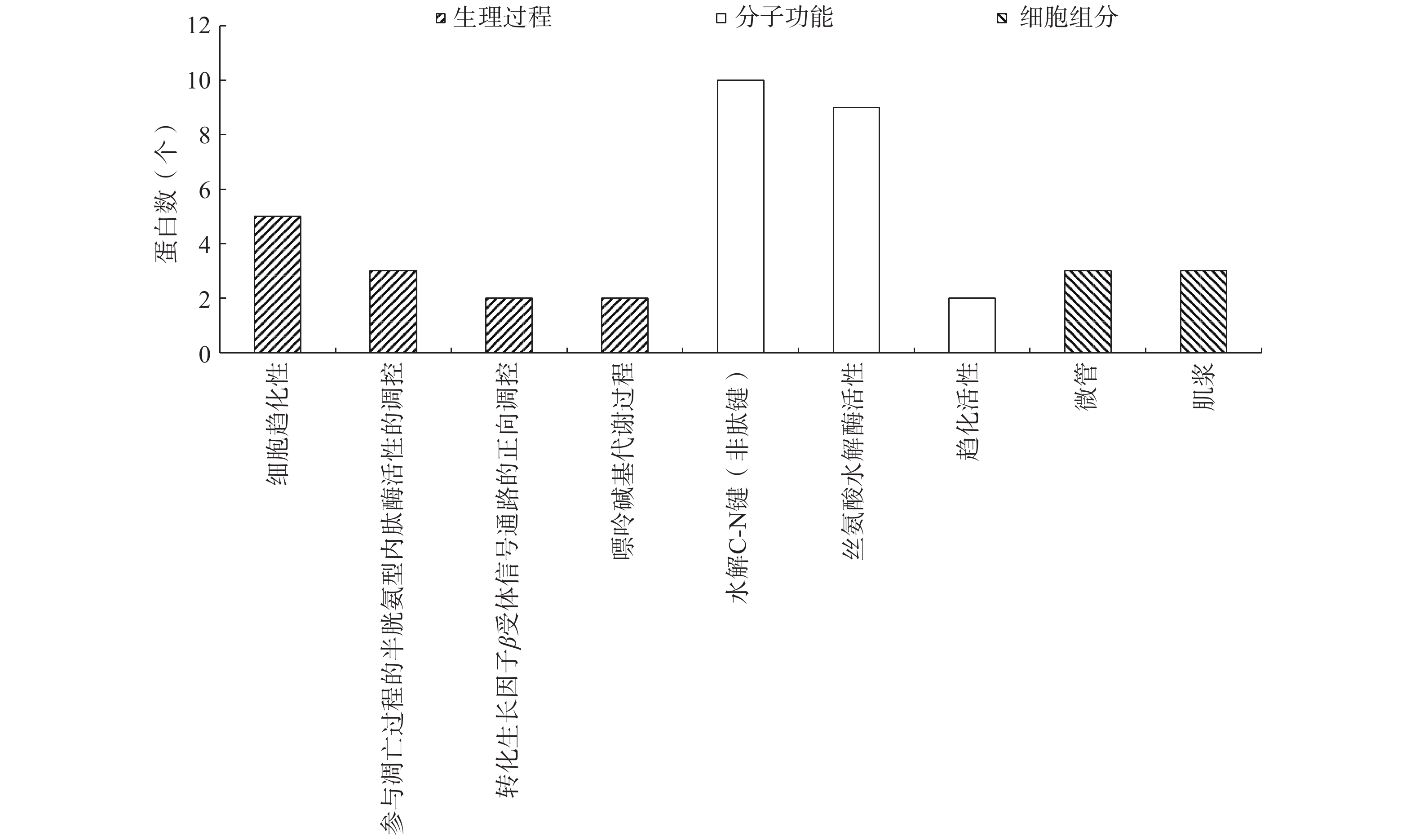
对参与的生理过程、具有的分子功能和所处的细胞组分对小鼠血清低丰度表达蛋白质进行分类,再通过Fisher精确检验方法对小鼠血清低丰度表达的26个差异表达蛋白进行GO功能富集分析,结果如图6和表5所示。结果表明,在小鼠血清低丰度表达蛋白质中,与ADB-M组相比,ADB-T组有5、3、2、2种显著表达的蛋白质分别参与了细胞趋化性、凋亡过程中半胱氨酸型肽链内切酶活性、转化生长因子β受体信号通路正向调节、嘌呤碱基代谢过程等4个生物过程,其富集度分别为0.227、0.333、0.667、0.667;而具有对C-N键(非肽键)作用的水解酶活性、丝氨酸水解酶活性、趋化活性等3个生物功能的蛋白质分别有10、9、2种表达显著;存在于微管和胞浆组分中的差异表达蛋白各有3个,其富集度分别为0.6和0.375。
![]() 图 6 差异表达蛋白GO功能富集结果Figure 6. Enriched GO terms of proteins with significant differences表 5 GO功能富集度Table 5. Richfactor of enriched GO terms
图 6 差异表达蛋白GO功能富集结果Figure 6. Enriched GO terms of proteins with significant differences表 5 GO功能富集度Table 5. Richfactor of enriched GO termsGO功能分类 细胞
趋化性参与凋亡过程的半胱氨型
内肽酶活性的调控转化生长因子β受体信号
通路的正向调控嘌呤碱基
代谢过程水解C-N键
(非肽键)丝氨酸水解
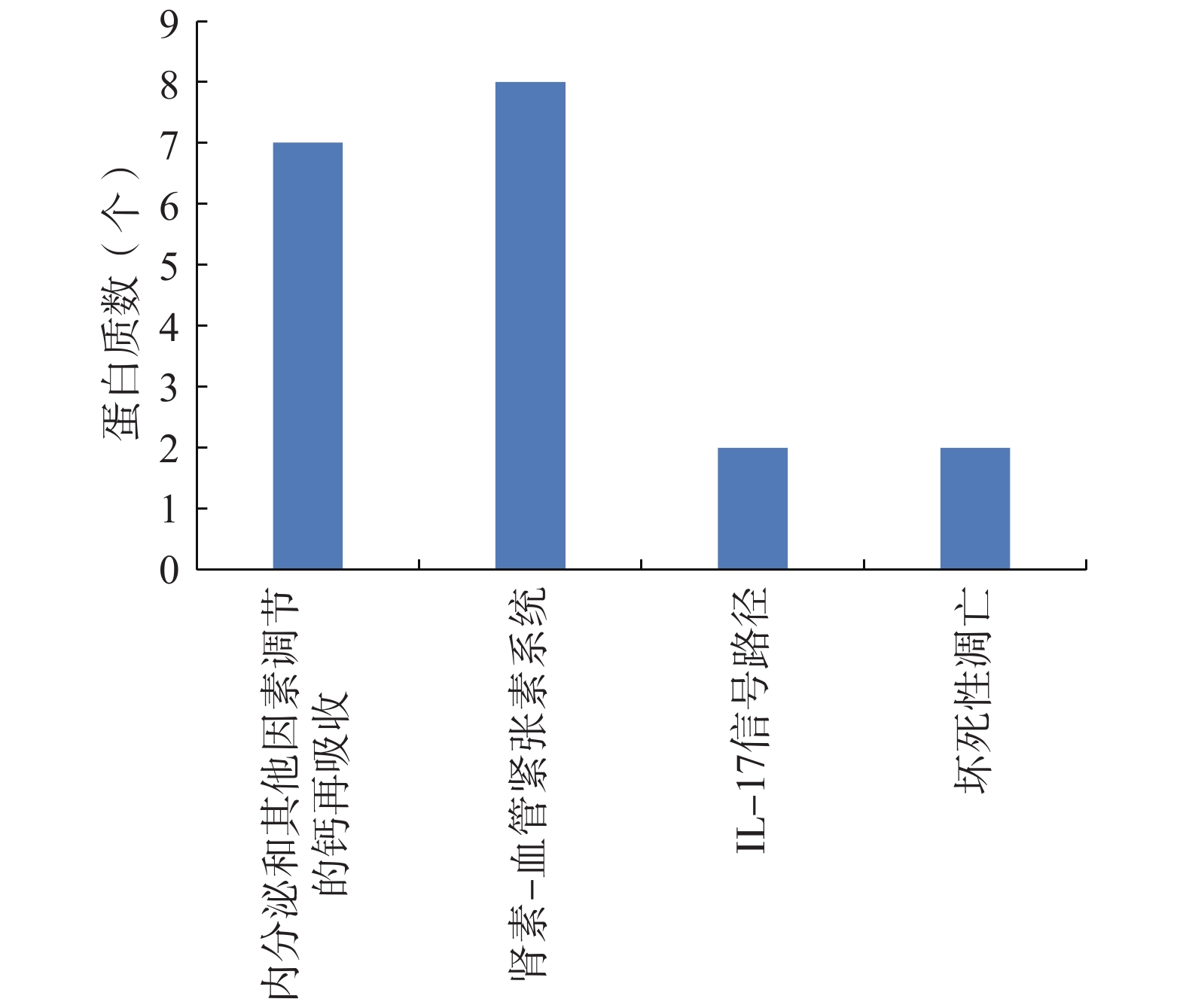
酶活性趋化活性 微管 肌浆 富集度 0.227 0.333 0.667 0.667 0.476 0.2 0.5 0.6 0.375 生物体内蛋白质在行使其功能时,受多方面因素调节,包括与其它蛋白质之间的协调作用。这些相互作用构成代谢通路。通过观察大鲵活性肽治疗前后蛋白质在KEGG通路中表达的变化,可以在蛋白质层面获得大鲵活性肽对治疗小鼠衰老的机理。如图7所示,通过Fisher精确检验方法对差异表达蛋白质进行KEGG通路富集分析,结果表明,内分泌调节的及其它因素调节的钙再吸收,肾素-血管紧张素系统,IL-17信号通路和坏死性凋亡等重要通路发生了显著变化。
3. 讨论
衰老是多细胞生命体生长发育成熟后在遗传、环境与生活方式等多种因素共同作用下引起生理机能逐步退行性下降的过程,小鼠衰老时表观学上为毛色灰暗,体重增长缓慢,精神萎靡等[19]。已有文献表明,机体衰老与体内自由基的量有关[9,13],抗衰老活性物质一般具有清除自由基活性。具有抗衰老活性的生物活性物质可令代谢紊乱小鼠代谢趋向正常化,其体内相关抗氧化酶和还原性物质含量得到改善,例如抗氧化蛋白[25]、超氧化物歧化酶[26]等,也可以降低衰老小鼠MDA含量。
在本研究中,通过对小鼠连续6周注射D-半乳糖后,模型组与灌胃大鲵活性肽组相比,小鼠血清低丰度蛋白质表达差异的有26个,包括四氢叶酸合成酶、糖原磷酸化酶和黄嘌呤脱氢酶等蛋白质。四氢叶酸合成酶是叶酸代谢途径中的关键酶,在一碳单位转移过程起作用,主要参与核酸代谢、蛋白质与核酸的甲基化与修复反应[27],为生物进行正常生长代谢、细胞损伤修复提供了必要的物质基础。本研究中衰老小鼠接受大鲵活性肽后,胞质四氢叶酸合成酶表达量上调,有利于D-半乳糖致衰老小鼠恢复正常。糖原磷酸化酶是糖原分解为1-磷酸葡萄糖的限速酶,其量的增加或降低影响生物的供能、高分子化合物合成代谢重要中间化合物来源[28]。本研究中的衰老小鼠经大鲵活性肽的调理后,糖原磷酸化酶表达量上升,有利于衰老小鼠恢复血糖浓度,进入正常生长状态。黄嘌呤脱氢酶参与嘌呤代谢,催化产生高氧化能力的过氧化物,使得机体易受氧化损伤[29-31]。灌胃大鲵活性肽后,衰老小鼠黄嘌呤脱氢酶表达量降低,在一定程度上可以减少过氧化物的含量,使得衰老小鼠脏器免受过氧化物的攻击。因此,对D-半乳糖致衰小鼠灌胃大鲵生物活性肽,可以改善小鼠体内T-AOC、SOD和MDA,并使多个代谢关键酶的表达量增加或减少,改善小鼠衰老状态,因此本研究所用大鲵生物活性肽具有抗衰老活性。
4. 结论
本研究通过D-半乳糖致小鼠衰老实验,发现大鲵活性肽可以使衰老小鼠体重和脾指数恢复增加,并引起SOD和T-AOC升高,降低肝指数和MDA含量,增强机体免疫,改善衰老小鼠紊乱的生化指标。衰老小鼠血清低丰度蛋白质中胞质四氢叶酸合成酶和肌糖原磷酸化酶表达量增加,黄嘌呤脱氢酶表达量减少,这些蛋白质表达量的变化,可以降低机体中自由基,延缓衰老进程。这些结果表明,分量为1400~2000 Da的大鲵活性肽具有抗小鼠衰老作用。
-
![]()
图 1 大鲵活性肽对小鼠体质量的影响
注:同组不同小写字母,表示为同组不同时间差异显著(P<0.05);同组相同小写字母,表示为同组不同时间差异不显著(P>0.05)。
Figure 1. Effect of giant salamander bioactive peptides on body weight of mice
![]()
图 2 小鼠血清样本平行性
注:A:小鼠血清蛋白SDS-PAGE,其中Marker为标准蛋白,ADB-M-1、ADB-M-2、ADB-M-3为模型组去除高丰度蛋白血清三个平行样,ADB-T2-1、ADB-T2-2、ADB-T2-3为中剂量组去除高丰度蛋白血清三个平行样,ADB-T2-3-原为中剂量组血清;B:去除高丰度蛋白的模型组小鼠血清蛋白质谱图;C:去除高丰度蛋白的中剂量组小鼠血清蛋白质谱图。
Figure 2. Samples parallelism of proteins in serum in mice
![]()
图 6 差异表达蛋白GO功能富集结果
Figure 6. Enriched GO terms of proteins with significant differences
表 1 大鲵活性肽对小鼠脏器指数的影响
Table 1 Effect of giant salamander bioactive peptides on organ indexes for each group of mice
分组 脑指数
(mg/10 g)肝脏指数
(mg/10 g)胸腺指数
(mg/10 g)脾指数
(mg/10 g)正常组 98.8±26.4 393.5±39.9 20.5±12.2 25.2±8.3 模型组 99.4±24.0 462.3±15.8* 21.9±9.6 31.9±8.8 低剂量组 91.1±16.0 399.9±55.2## 24.4±7.5 30.9±4.5 中剂量组 94.1±15.5 454.4±25.6* 22.2±4.3 32.9±8.0* 高剂量组 87.9±17.2 415.2±30.6## 23.4±14.0 28.6±6.5 注:*表示与正常组相比,差异显著(P<0.05);**表示与正常组相比,差异极显著(P<0.01);#表示与模型组相比,差异显著(P<0.05);##表示与模型组相比,差异极显著(P<0.01);表2同。  下载: 导出CSV
下载: 导出CSV
表 2 大鲵活性肽对小鼠生化指标的影响
Table 2 Effect of giant salamander bioactive peptides on the levels of serum markers in SPF mice
分组 MDA(nmol/mL) SOD活力(U/mL) T-AOC(mmol/L) 正常组 10.58±1.88 157.26±16.15 0.58±0.04 模型组 16.70±2.60** 119.88±17.49** 0.48±0.05** 低剂量组 14.86±2.10** 131.35±13.99** 0.50±0.05** 中剂量组 11.90±2.60## 141.16±16.93*## 0.54±0.05*# 高剂量组 12.08±2.27## 138.51±15.59*# 0.54±0.04*#
下载: 导出CSV
表 3 上调的差异表达蛋白质
Table 3 Up-regulated proteins with significant differences
存取编号 蛋白质 分子量(kDa) 中剂量组表达量/
模型组表达量t检验P值 pI 参与代谢途径 P07628 激肽释放酶1相关肽酶b8 28.531 4.49 1.43×10−4 8.28 内分泌和其它因素调节的钙再吸收
肾素-血管紧张素系统Q9JM71 激肽释放酶1相关肽酶b27 28.742 4.18 2.75×10−5 8.86 P00796 肾素-2 44.282 3.41 3.49×10−5 6.01 肾素-血管紧张素系统/肾素分泌 Q61759 激肽释放酶1相关肽酶b21 28.69 2.93 1.80×10−4 7.05 内分泌和其它因素调节的钙再吸收
肾素-血管紧张素系统Q922D8 胞质四氢叶酸合成酶 101.2 2.78 1.24×10−3 6.7 叶酸一碳库 P15946 激肽释放酶1相关肽酶b11 28.727 2.70 2.15×10−5 6.69 内分泌和其它因素调节的钙再吸收
肾素-血管紧张素系统P00756 激肽释放酶1相关肽酶b3 28.998 2.54 1.00×10−4 6.37 内分泌和其它因素调节的钙再吸收
肾素-血管紧张素系统P15948 激肽释放酶1相关肽酶b22 28.384 2.52 5.15×10−5 6.2 内分泌和其它因素调节的钙再吸收
肾素-血管紧张素系统P15949 激肽释放酶1相关肽酶b9 28.9 2.45 1.54×10−4 7.56 内分泌和其它因素调节的钙再吸收
肾素-血管紧张素系统P04071 激肽释放酶1相关肽酶b16 28.722 2.18 2.54×10−6 5.4 P36369 激肽释放酶1相关肽酶b26 28.463 2.12 2.06×10−4 6.41 内分泌和其它因素调节的钙再吸收
肾素-血管紧张素系统P28666 鼠球蛋白-2 162.38 2.12 8.12×10−7 6.29 Q9WUB3 肌糖原磷酸化酶 97.285 2.05 3.37×10−5 6.65 胰岛素信号通路/淀粉和蔗糖代谢/坏死性凋亡
胰岛素抵抗/胰高血糖素信号通路注:中剂量组表达量/模型组表达量-表示中剂量治疗组和模型组蛋白质表达量平均值的比值;表4同。
下载: 导出CSV
表 4 下调的差异表达蛋白质
Table 4 Down-regulated proteins with significant differences
存取编号 蛋白质 分子量(kDa) 中剂量组表达量/模型组表达量 t检验P值 pI 参与代谢途径 P32261 抗凝血酶Ⅲ 52.003 0.49 1.16×10−4 6.1 补体和凝血级联反应 Q00519 黄嘌呤脱氢酶/氧化酶 146.56 0.49 3.26×10−2 7.62 过氧化物酶体/药物代谢-其他酶/嘌呤代谢/咖啡因代谢 Q61805 脂多糖结合蛋白 53.055 0.48 1.46×10−3 8.64 NF-κB信号通路/结核/Toll样受体信号通路/沙门氏菌感染 P35441 血小板反应蛋白-1 129.65 0.48 4.09×10−6 4.72 P13634 碳酸酐酶1 28.33 0.48 9.12×10−3 6.44 氮代谢 P68373 微管蛋白α-1C链 49.909 0.44 5.26×10−4 4.96 吞噬体/紧密连接/缝隙连接/凋亡 P11499 热休克蛋白hsp90-β 83.28 0.42 4.46×10−2 4.96 Th17细胞分化/坏死下垂/雌激素信号通路/内质网蛋白质处理/PI3K-Akt信号通路/抗原处理和提呈/IL-17信号通路/NOD样受体信号通路/癌症/孕酮介导的卵母细胞成熟/前列腺癌/流体剪切应力和动脉粥样硬化的途径 P07743 BPI折叠包含家族A成员2 24.753 0.33 9.06×10−5 4.87 Q71KU9 纤维蛋白原样蛋白1 36.439 0.28 4.40×10−4 5.48 P31725 蛋白质S100-A9 13.049 0.27 4.49×10−3 6.65 IL-17信号通路 P05366 血清淀粉样蛋白A-1 13.77 0.07 2.30×10−5 6.5 P05367 血清淀粉样蛋白A-2 13.622 0.02 8.21×10−5 6.4 P02089 血红蛋白β-2亚基 15.878 0.00 9.12×10−6 7.85
下载: 导出CSV
表 5 GO功能富集度
Table 5 Richfactor of enriched GO terms
GO功能分类 细胞
趋化性参与凋亡过程的半胱氨型
内肽酶活性的调控转化生长因子β受体信号
通路的正向调控嘌呤碱基
代谢过程水解C-N键
(非肽键)丝氨酸水解
酶活性趋化活性 微管 肌浆 富集度 0.227 0.333 0.667 0.667 0.476 0.2 0.5 0.6 0.375
下载: 导出CSV
-
[1] 罗庆华, 刘英, 张立云. 张家界市大鲵资源保护·增殖现状与对策[J]. 安徽农业科学,2009,.37(19):9023−9025, 9052. [2] 李时珍. 本草纲目[M]. 北京: 人民卫生出版社, 2005. [3] 谢宗万. 全国中草药汇编[M]. 北京: 人民卫生出版社, 1978. [4] 李莉, 王锡昌, 刘源. 中国养殖大鲵的食用、药用价值及其开发利用研究进展[J]. 食品工业科技,2012,33(9):454−458. [5] 纪钱萍, 李伟, 佟长青. 大鲵活性肽牡蛎多糖复合粉体外ACE抑制作用的研究[J]. 农产品加工,2020(20):29−31. [6] 贺屹潮. 大鲵肉酶解肽制备及抗氧化与免疫调节活性研究[D]. 汉中: 陕西理工大学, 2020. [7] 张佳婵, 王昌涛, 李萌, 等. 大鲵活性肽酶法制备工艺优化及抗氧化性分析[J]. 食品工业科技,2016,37(12):217−221, 242. [8] 李素云, 王立芹, 郑稼琳, 等. 自由基与衰老的研究进展[J]. 中国老年学杂志, 2007, 27(20): 2046−2048. [9] 王荣, 杨宽, 陈春妮, 等. 亚麻籽提取物对D-半乳糖致衰老小鼠的抗氧化保护机制研究[J]. 中国油脂,2019,44(8):92−95. [10] Qu Z, Zhang J, Yang H, et al. Protective effect of tetrahydropalmatine against d-galactose induced memory impairment in rat[J]. Physiology & Behavior,2016,154:114−125.
[11] Gao J, He H, Jiang W, et al. Salidroside ameliorates cognitive impairment in a d-galactose-induced rat model of Alzheimer’s disease[J]. Behavioural Brain Research,2015,293:27−33. doi: 10.1016/j.bbr.2015.06.045
[12] 王颖, 王欣卉, 徐炳政, 等. 芸豆芽菜多酚对D-半乳糖致小鼠机体氧化及肾脏损伤的修复作用[J]. 食品科学,2017,38(13):219−223. doi: 10.7506/spkx1002-6630-201713036 [13] 黄杰, 董照瀛, 许梦雄, 等. D-半乳糖致小鼠胰腺损伤[J]. 基础医学与临床,2017,37(7):912−917. [14] Gong Y-S, Guo J, Hu K, et al. Ameliorative effect of lotus seedpod proanthocyanidins on cognitive impairment and brain aging induced by d-galactose[J]. Experimental Gerontology,2016,74:21−28. doi: 10.1016/j.exger.2015.11.020
[15] 王爱萍, 史明仪, 费文勇, 等. 银杏外种皮内酯对D-半乳糖致脑衰老小鼠的作用[J]. 中国中医基础医学杂志,2005(3):189−191. doi: 10.3969/j.issn.1006-3250.2005.03.011 [16] 王文莉, 张伟, 于新莹, 等. 大鲵肉酶解产物的制备及其抗氧化性的研究[J]. 河北渔业,2012(9):1−4. [17] 付静, 陈德经, 曹米娜. 大鲵多肽制备工艺的研究[J]. 食品科技,2012,37(2):66−68, 72. [18] Wiśniewski J R, Zougman A, Nagaraj N, et al. Universal sample preparation method for proteome analysis[J]. Nature Methods,2009,6(5):359−362. doi: 10.1038/nmeth.1322
[19] 翟兴月, 王庆辉, 赵冠华, 等. 非标记定量蛋白质组方法分析鲟鱼肽抗D-半乳糖导致的小鼠衰老作用的研究[J]. 食品工业科技,2019,40(3):290−295. [20] Cox J, Hein M Y, Luber C A, et al. Accurate proteome-wide label-free quantification by delayed normalization and maximal peptide ratio extraction, termed MaxLFQ[J]. Molecular & Cellular Proteomics,2014,13(9):2513−2526.
[21] Cox J, Mann M. MaxQuant enables high peptide identification rates, individualized p.p.b.-range mass accuracies and proteome-wide protein quantification[J]. Nature Biotechnology,2008,26(12):1367−1372. doi: 10.1038/nbt.1511
[22] Zhu Y, Xu H, Chen H, et al. Proteomic analysis of solid pseudopapillary tumor of the pancreas reveals dysfunction of the endoplasmic reticulum protein processing pathway[J]. Molecular & Cellular Proteomics,2014,13(10):2593−2603.
[23] Gotz S, Garcia-Gomez J M, Terol J, et al. High-throughput functional annotation and data mining with the Blast2GO suite[J]. Nucleic Acids Research,2008,36(10):3420−3435. doi: 10.1093/nar/gkn176
[24] Moriya Y, Itoh M, Okuda S, et al. KAAS: An automatic genome annotation and pathway reconstruction server[J]. Nucleic Acids Research,2007,35(Web Server issue):W182−W185.
[25] 车晓宁. 有氧运动对衰老的大鼠脑组织抗氧化蛋白的影响[J]. 职业与健康,2014,30(13):1782−1784. [26] 韦忠建, 陆碧琼, 胡江平. 不同负荷的间歇性游泳运动对衰老小鼠腓肠肌丙二醛含量和超氧化物歧化酶活性的影响[J]. 中国老年学杂志,2019,39(15):3781−3783. doi: 10.3969/j.issn.1005-9202.2019.15.057 [27] Chen M, Zhai J, Liu Y, et al. Molecular cloning and characterization of C1 tetrahydrofolate (C1-THF) synthase in Bombyx mori, silkworm[J]. Gene,2018,663:25−33. doi: 10.1016/j.gene.2018.04.037
[28] Bai Y, Li X, Zhang D, et al. Effects of phosphorylation on the activity of glycogen phosphorylase in mutton during incubation at 4 degrees C in vitro[J]. Food Chemistry,2020,313:126162. doi: 10.1016/j.foodchem.2020.126162
[29] Chen Y, Li Yan, Chao H, et al. Molecular cloning and characterisation of a novel xanthine oxidase from Cellulosimicrobium cellulans ATCC21606[J]. Process Biochemistry,2020,91:65−72. doi: 10.1016/j.procbio.2019.11.033
[30] Al-Shehri S S, Duley J A, Bansal N. Xanthine oxidase-lactoperoxidase system and innate immunity: Biochemical actions and physiological roles[J]. Redox Biology,2020,34:101524. doi: 10.1016/j.redox.2020.101524
[31] Monika, Sharma N K, Thakur N, et al. Xanthine oxidase of Acinetobacter calcoaceticus RL2-M4: Production, purification and characterization[J]. Protein Expression and Purification,2019,160:36−44. doi: 10.1016/j.pep.2019.03.014

 下载:
下载:







计量
- 文章访问数: 319
- HTML全文浏览量: 88
- PDF下载量: 28
