


Study on Serine Proteinase in the Muscle of Yesso Scallops (Mizuhopecten yessoensis)
-
摘要: 为了探究虾夷扇贝肌肉冷藏过程中内源蛋白酶对其质构的影响,本文对虾夷扇贝肌肉在4 ℃下冷藏7 d过程中肌肉质构及蛋白变化进行测定,结果表明,整个冷藏过程中扇贝肌肉硬度、弹性、咀嚼性和粘性整体呈下降趋势。SDS-PAGE分析显示肌肉蛋白在第3 d开始出现明显降解。在内源酶活力方面,丝氨酸蛋白酶(Serine proteinase,SP)活力在冷藏2 d后开始急剧下降。通过硫酸铵盐析、离子交换、凝胶过滤和疏水柱层析等从虾夷扇贝肌肉中获得高度纯化的丝氨酸蛋白酶并对其酶学性质进行研究。SDS-PAGE和明胶酶谱结果表明,SP在天然状态下主要以分子量约为52 kDa的二聚体形式存在。SP的最适pH为9.0,最适温度为37 ℃。SP特异性水解羧基侧P1位含有精氨酸或赖氨酸残基的底物。丝氨酸蛋白酶特异性抑制剂PMSF、Leupeptin、Pefabloc SC、Benzamidine分别能抑制其97%、98%、90%和85%的酶活力;金属离子Fe2+、Zn2+、Cu2+也能明显抑制SP的酶活力。37 ℃下SP可有效降解扇贝肌原纤维蛋白,为揭示SP对扇贝肌肉蛋白的品质影响提供参考。Abstract: To investigate the effect of endogenous protease on the texture of Yesso scallop muscle during cold storage, the texture and protein change related indexes of Yesso scallop muscle was measured during 4 ℃ cold storage 7 d with a purpose to identify the functions of proteinases. The results showed that the hardness, springiness, chewiness and gumminess generally decreased during the whole cold storage process. SDS-PAGE analysis revealed that muscular proteins began to degrade obviously on the third day. In terms of enzyme activity, serine proteinase (SP) activity began to decrease sharply after 2 days of cold storage. Serine proteinase was purified from the muscle of Yesso scallop by ammonium sulfate precipitation, ion exchange chromatography, gel filtration and hydrophobic chromatography. SDS-PAGE and gelatin zymogram showed that in native condition SP was in homodimer form with molecular weight of 52 kDa. The optimum pH of SP was 9.0 and the optimum temperature was 37 ℃. SP specifically hydrolyzes substrates containing arginine or lysine residues at P1 position on the carboxyl side. Serine protease specific inhibitors including PMSF, Leupeptin, Pefabloc SC and Benzamidine could inhibit the activity of SP by 97%, 98%, 90% and 85%, respectively. Metal ions Fe2+, Zn2+ and Cu2+ could also significantly suppress the activity of SP. SP could effectively degrade scallop myofibrillar at 37 ℃, which delivered a reference for revealing the effect of SP on the quality of Yesso scallop muscle protein.
-
Keywords:
- Mizuhopecten yessoensis /
- serine proteinase /
- texture /
- purification /
- characterization /
- muscle
-
扇贝是我国主要经济贝类,2020年,全国扇贝养殖产量达175万吨[1]。扇贝的主要食用部位是闭壳肌,肉质细嫩、味道鲜美、营养丰富,具有很高的食用价值,深受消费者喜爱[2]。扇贝蛋白质含量丰富,脂肪、碳水化合物含量低,是优质的蛋白源。此外,还含有较丰富的维生素A、维生素B族、维生素E等[3-4]。
扇贝收获后的运输过程中,为了保持和延长其新鲜度和适口性,需要采取良好的冷藏手段[5]。对去壳后的扇贝闭壳肌而言,在冷藏过程中,其质地、风味等感官品质会不可避免发生下降,对产品的商品价值产生不利影响[6]。这主要是因为水产动物死后,维持肌肉结构特性的肌原纤维蛋白和结缔组织蛋白等结构蛋白发生变化,包括肌纤维断裂、保水性能降低等[7-9]。研究发现,由内源蛋白酶水解蛋白质引起的肌肉蛋白结构变化是影响水产动物蛋白质地下降的主要原因[10-12]。例如,罗氏沼虾在冷藏期间,丝氨酸蛋白酶的作用使其肌肉软化、品质下降[13]。鱼类肌肉中肌原纤维结合型丝氨酸蛋白酶对肌球蛋白重链的降解引起鱼糜制品的凝胶劣化[14-15]。扇贝中结构蛋白的完整性与肌肉质地指标密切相关。最近,有学者对海湾扇贝的研究表明,丝氨酸蛋白酶可以同时降解肌原纤维蛋白和结缔组织蛋白,进而改变肌肉微观结构,使其持水力降低,最终引起扇贝肌肉质地的劣化[16]。
丝氨酸蛋白酶(Serine proteinase,SP)是蛋白酶家族中的重要一员,作为内肽酶,SP特异性作用于大分子蛋白质中肽键C端的精氨酸或赖氨酸残基,使之断裂成为小分子蛋白质或多肽。该酶的关键活性位点通常是一组含有丝氨酸、组氨酸和天冬氨酸的催化三联体。在无脊椎动物中,丝氨酸蛋白酶及其同源物广泛存在并被证明在生物体的各种生理生化过程发挥重要作用[17-19]。作为主要经济贝类,扇贝在冷藏加工过程中的品质保持尤为重要。但是,对影响扇贝品质的丝氨酸蛋白酶的分离纯化和性质研究尚未有报道。因此,本研究拟以虾夷扇贝为研究对象,探究其闭壳肌在冷藏过程中的质地变化,并分离纯化出丝氨酸蛋白酶,对其酶学性质进行研究,以期为贝类在保鲜贮藏和加工过程的品质保持提供一定的理论参考。
1. 材料与方法
1.1 材料与仪器
原产河北秦皇岛的鲜活虾夷扇贝(Mizuhopecten yessoensis) 厦门市集美区新华都商场,去壳后贝柱个体重量(15±0.5)g;DEAE-Sepharose、Superdex 200、Phenyl-Sepharose 美国GE Healthcare公司;Boc-Phe-Ser-Arg-MCA等各类荧光合成底物 日本Peptide Institute公司;蛋白酶抑制剂:PMSF、EDTA、EGTA、1,10-phenanthroline 美国Sigma公司;E-64、Leupeptin、Triton X-100 美国Amresco公司;Pefabloc 德国SC Roche公司;牛明胶 美国Genebase公司。
Avanti J-26S XP高速冷冻离心机 美国Beckman公司;Centrifuge 5417小型冷冻离心机 德国Eppendorf公司;UB-7 Starter 3100 pH计 美国Ohaus公司;UPC900 AKTA 蛋白纯化系统 美国GE Healthcare公司;FP-8200荧光分光光度计 日本Jasco公司;TA new plus质构仪 美国Isenso公司;WB-10L1数显恒温水浴锅 德国Memmert公司;PT-2100组织破碎机 瑞士Kinematica公司;G-BOX凝胶成像仪 英国Syngene公司;Mini-PROTEAN垂直电泳槽 美国Bio-Rad公司;ND-1000微量蛋白核酸测定仪 美国Nano Drop公司。
1.2 实验方法
1.2.1 冷藏过程中肌肉质构相关指数的测定
参照游银川[20]的方法稍加改动,具体方法为新鲜虾夷扇贝,去壳去内脏团,取其肌肉,浸泡于0.2% NaN3溶液中5 min,沥干后用自封袋真空密封,储藏于4 ℃,每隔24 h取样直至一周。
1.2.1.1 冷藏过程中肌肉质构参数(TPA)的测定
参照游银川[20]的方法稍加改动,具体方法为将虾夷扇贝贝柱肌肉裁剪成高(15.0±0.1)mm、直径(35.0±0.2)mm的圆柱体,采用TA new plus质构仪,通过P/36R探头,对样品进行全质构分析(TPA)。压缩变形模量为25%,探头速度为1.0 mm/s,每个样品分别进行2次循环压缩分析,循环之间的松弛时间为5 s。得到硬度、咀嚼性、粘性和弹性等质构参数。每组数据用4个贝柱独立测定,取其平均值。
1.2.1.2 冷藏过程肌肉全蛋白的降解
参照游银川[20]的方法稍加改动,具体方法为取2 g扇贝肌肉,加入4倍含0.15 mol/L NaCl的20 mmol/L Tris-HCl(pH7.5)缓冲液,组织捣碎后离心(3000×g,4 ℃,5 min),取上清,即为肌肉全蛋白。样品加入SDS化缓冲液,振荡混匀,95 ℃加热10 min后4 ℃保存。采用SDS-PAGE对肌肉全蛋白降解进行分析。
1.2.1.3 冷藏过程丝氨酸蛋白酶酶活力的测定方法
参照游银川[20]的方法稍加改动,具体方法为取2 g扇贝肌肉,加入4倍体积的20 mmol/L Tris-HCl(pH7.5)缓冲液,组织捣碎后离心(12000×g,4 ℃,15 min),上清即为测定内源酶粗酶活力的样品。
酶活力测定参照林怡晨等[21]的方法稍加改动,具体方法为,以Boc-Phe-Ser-Arg-MCA为丝氨酸蛋白酶底物测定酶活力。取50 μL粗酶液置于900 μL 20 mmol/L Tris-HCl(pH9.0)缓冲液中,加入50 μL浓度为10 μmol/L的底物后,振荡混匀后在37 ℃下孵育10 min,立即加入1.5 mL终止液(甲醇:正丁醇:蒸馏水=35:30:35,v/v/v)终止反应。用荧光分光光度计在激发波长380 nm,发射波长450 nm下测定释放的7-氨基-4-甲基香豆素(7-amino-4-methylcoumarin,AMC)的荧光强度。酶活力单位(U)定义为每分钟释放1 nmol AMC所需要的酶量。以同样的反应体系但不加酶液(缓冲液替代)的样品为空白对照组。
1.2.2 丝氨酸蛋白酶的分离纯化
所有纯化过程均在4 ℃或冰上进行。将250 g虾夷扇贝肌肉加入4倍体积的20 mmol/L Tris-HCl(pH9.0)缓冲液,组织捣碎后离心(12000×g,4 ℃,20 min),上清进行30%~60%硫酸铵盐析,离心后将沉淀用少量的缓冲液溶解,用20 mmol/L Tris-HCl(pH9.0)缓冲液充分透析后离心(12000×g,4 ℃,20 min),上清液用4层纱布过滤后上样于DEAE-Sepharose 离子交换层析柱。先用20 mmol/L Tris-HCl(pH9.0)缓冲液流洗,再用含0.5 mol/L NaCl的缓冲液进行线性洗脱,收集目的蛋白组分,用10 kDa的超滤浓缩膜浓缩至3 mL。随后上样于Superdex 200凝胶过滤柱,用20 mmol/L Tris-HCl(pH9.0)缓冲液洗脱,收集目的蛋白组分加入(NH4)2SO4至终浓度为1.0 mol/L,然后上样于Phenyl-Sepharose疏水层析柱(5 mL)。用含1.0 mol/L(NH4)2SO4的缓冲液洗脱,收集目的蛋白组分,用20 mmol/L Tris-HCl(pH9.0)缓冲液充分透析,再次上样于DEAE-Sepharose 柱层析(5 mL),洗脱收集目的蛋白组分即为高度纯化的丝氨酸蛋白酶。
1.2.3 SDS-PAGE和明胶酶谱分析
SDS-PAGE凝胶为12%分离胶和5%浓缩胶。明胶酶谱参考麻金花等[22]的方法,略作修改,分离胶中用 0.5 mL浓度为10 mg/mL明胶替换 0.5 mL去离子水,其余与SDS-PAGE胶的制备相同。4 ℃条件下电泳结束,用2.5% Triton X-100孵育20 min,洗去SDS。然后置于20 mmol/L Tris-HCl(pH9.0)缓冲液中,于37 ℃下孵育14 h。考马斯亮蓝R-250染色后,凝胶未着色的亮带表示明胶被蛋白酶降解,鉴定蛋白酶对明胶的分解活性。
1.2.4 扇贝肌肉丝氨酸蛋白酶的酶学性质分析
1.2.4.1 最适温度及热稳定性分析
测定SP的最适温度,以Boc-Phe-Ser-Arg-MCA为底物,在10、20、30、35、37、40、50、60、70 ℃等温度下测定酶活力。将酶活力最高所对应温度的相对活力定义为100%,以此来判定其它温度下的相对酶活力。
测定SP的热稳定性,将SP于上述不同温度下孵育30 min并冷却至室温后,在37 ℃下测定相对酶活力。
1.2.4.2 最适pH及pH稳定性分析
测定SP的最适pH,以Boc-Phe-Ser-Arg-MCA为底物,在不同pH(4.0、5.0、6.0、7.0、8.0、9.0、10.0、11.0)下测定酶活力。将酶活力最高pH下的相对活力定义为100%,以此来判定其他pH下的相对酶活力。
测定SP的pH稳定性,将SP置于不同pH(4.0、5.0、6.0、7.0、8.0、9.0、10.0、11.0)缓冲液中,4 ℃孵育4 h后,在20 mmol/L Tris-HCl(pH9.0)缓冲液中测定酶活力。
各pH所用缓冲液:pH4.0~5.0 为20 mmol/L HAc-NaAc 缓冲液、pH6.0为20 mmol/L PBS缓冲液、pH7.0~9.0 为20 mmol/L Tris-HCl 缓冲液、pH10.0~11.0为20 mmol/L Na2CO3-NaHCO3缓冲液。
1.2.4.3 底物特异性分析
参照林怡晨等[21]的方法稍加改动,具体方法为采用Boc-Phe-Ser-Arg-MCA等各类荧光合成底物,参照酶活力测定方法来分析SP对不同底物的水解能力。以荧光值最高的底物为空白对照。
1.2.4.4 蛋白酶抑制剂对SP活力的影响
参照林怡晨等[21]的方法稍加改动,具体方法为将PMSF,EDTA,EGTA,1,10-phenanthroline等蛋白酶抑制剂与SP混匀后4 ℃下孵育30 min,测定剩余酶活力。以不加抑制剂的样品为对照。
1.2.4.5 金属离子对SP酶活力的影响
在20 mmol/L Tris-HCl(pH9.0)缓冲液中,加入50 μL的酶液,加入不同金属离子(Mn2+、Mg2+、Ca2+、Cu2+、Fe2+、Zn2+、Ba2+)至终浓度分别为0.01、0.05和0.1 mmol/L,混匀后室温下孵育30 min,在37 ℃下进行酶活力测定。以不加金属离子的样品为对照。
1.2.5 丝氨酸蛋白酶对虾夷扇贝肌原纤维的分解
参考Cao等[23]的方法提取肌原纤维蛋白。具体方法为:取10 g扇贝肌肉,加入10倍体积20 mmol/L Tris-HCl(pH7.5)缓冲液,组织捣碎后离心(8000×g,4 ℃,10 min),取沉淀重复该步骤4次后,将沉淀溶于含0.5 mol/L NaCl的20 mmol/L Tris-HCl(pH7.5)缓冲液,即为肌原纤维蛋白。
取上述制备的肌原纤维蛋白50 μL,溶于220 μL含0.5 mol/L NaCl的20 mmol/L Tris-HCl(pH7.5)缓冲液中,加入30 μL丝氨酸蛋白酶,37 ℃反应0、5、10、15、30、60、90、120 min,分别取出10 μL加入10 μL 4×SDS缓冲液混匀后,进行SDS-PAGE 分析。以不加酶的肌原纤维在37 ℃孵育120 min 为对照。
1.3 数据处理
采用SPSS 18.0软件进行数据分析,所有实验数据均重复3次进行测定。应用Origin 8.0软件进行绘图,Image J软件进行光密度分析。
2. 结果与分析
2.1 冷藏过程中肌肉质构相关指数的变化
2.1.1 冷藏过程中肌肉的质构参数(TPA)
对虾夷扇贝在冷藏过程中的质构变化进行测定,结果如表1所示。与新鲜样品相比,冷藏1 d,扇贝肌肉的硬度由670±42 gf增至736±52 gf,粘性321±2 gf增至372±17 gf,弹性0.34±0.07 gf增至0.37±0.04 gf,除咀嚼性外,都有增加。这与海湾扇贝[24]、鲷鱼[25]和虾[26]等有相同的变化趋势,推测这是由于虾夷扇贝肌肉冷藏1 d期间发生僵直所致。在此后的整个贮藏过程中,肌肉硬度、弹性、咀嚼性和粘性均不断下降,肉质劣化。水产动物肌肉在冷藏过程中的质构变化有类似之处,如鲑鱼(Salmo salar)在冷藏期间肌肉容易变软、失去弹性,硬度下降等现象[27]。
表 1 冷藏过程中虾夷扇贝肌肉质构参数变化Table 1. The change of texture parameters in Yesso callop muscle during cold storage时间 (d) 硬度 (gf) 咀嚼性(gf) 粘性(gf) 弹性(gf) 0 670±42b 255±18 a 321±2 b 0.34±0.07b 1 736±52a 176±11b 372±17a 0.37±0.04a 2 584±45c 152±8c 276±19c 0.33±0.02b 3 457±38d 132±4d 253±11d 0.32±0.03b 4 330±33e 126±5e 227±12e 0.31±0.02c 5 206±25f 115±2f 176±15f 0.28±0.01d 6 146±27g 102±3g 107±8g 0.27±0.03e 7 68±12h 93±4g 99±7g 0.25±0.02f 注:同列不同小写字母表示差异显著(P<0.05)。 2.1.2 冷藏过程中肌肉蛋白的自身降解
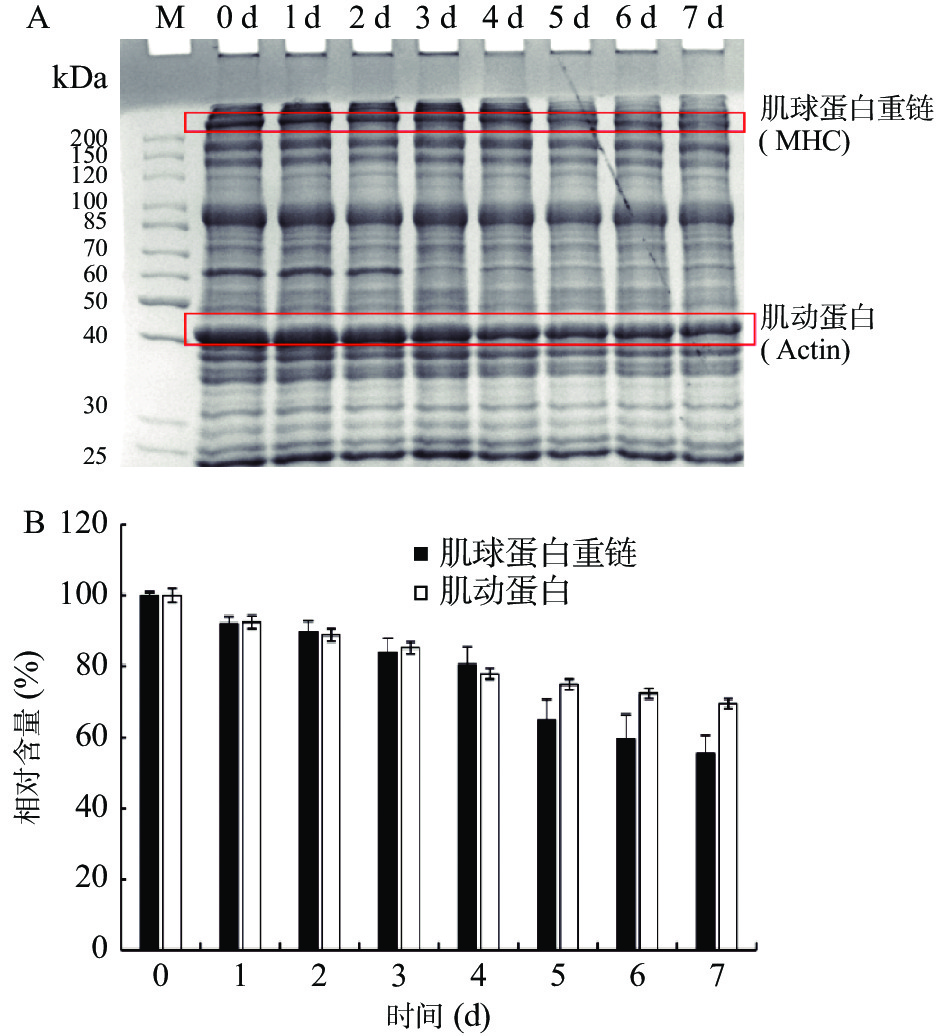
采用SDS-PAGE对虾夷扇贝肌肉全蛋白进行分析,以确定冷藏中蛋白的变化。肌原纤维蛋白是肌肉中的主要蛋白质[8]。随着冷藏时间的延长,全蛋白逐渐降解,观察到肌球蛋白重链(myosin heavy chain,MHC)(200 kDa)和肌动蛋白(Actin)(45 kDa)条带浓度逐渐减弱(图1A)。对其进行光密度分析密度,结果显示MHC和肌动蛋白的蛋白含量逐渐降低(图1B),表明冷藏过程中以肌球蛋白重链为代表的肌肉结构蛋白逐渐降解。有研究证明,肌球蛋白重链是肌原纤维蛋白中最容易被分解的蛋白[11,28]。此外,分子量为62 kDa的蛋白也在冷藏2 d后发生明显降解。分子量为100 kDa的副肌球蛋白是贝类的特征蛋白,该蛋白稳定性良好,在7 d的冷藏过程中几乎没有发生降解。水产动物结构蛋白在冷藏过程中随着时间的推移而降解是普遍现象,而内源性蛋白酶的作用是导致结构蛋白降解,肌肉质构下降的主要原因[29]。
![]() 图 1 虾夷扇贝冷藏过程中肌肉蛋白的自身降解(A)和光密度分析(B)注:M:标准蛋白。Figure 1. Protein degradation in Yesso scallop muscle during cold storage (A) and densitometry analysis (B)
图 1 虾夷扇贝冷藏过程中肌肉蛋白的自身降解(A)和光密度分析(B)注:M:标准蛋白。Figure 1. Protein degradation in Yesso scallop muscle during cold storage (A) and densitometry analysis (B)2.1.3 冷藏过程中丝氨酸蛋白酶酶活力水平的变化
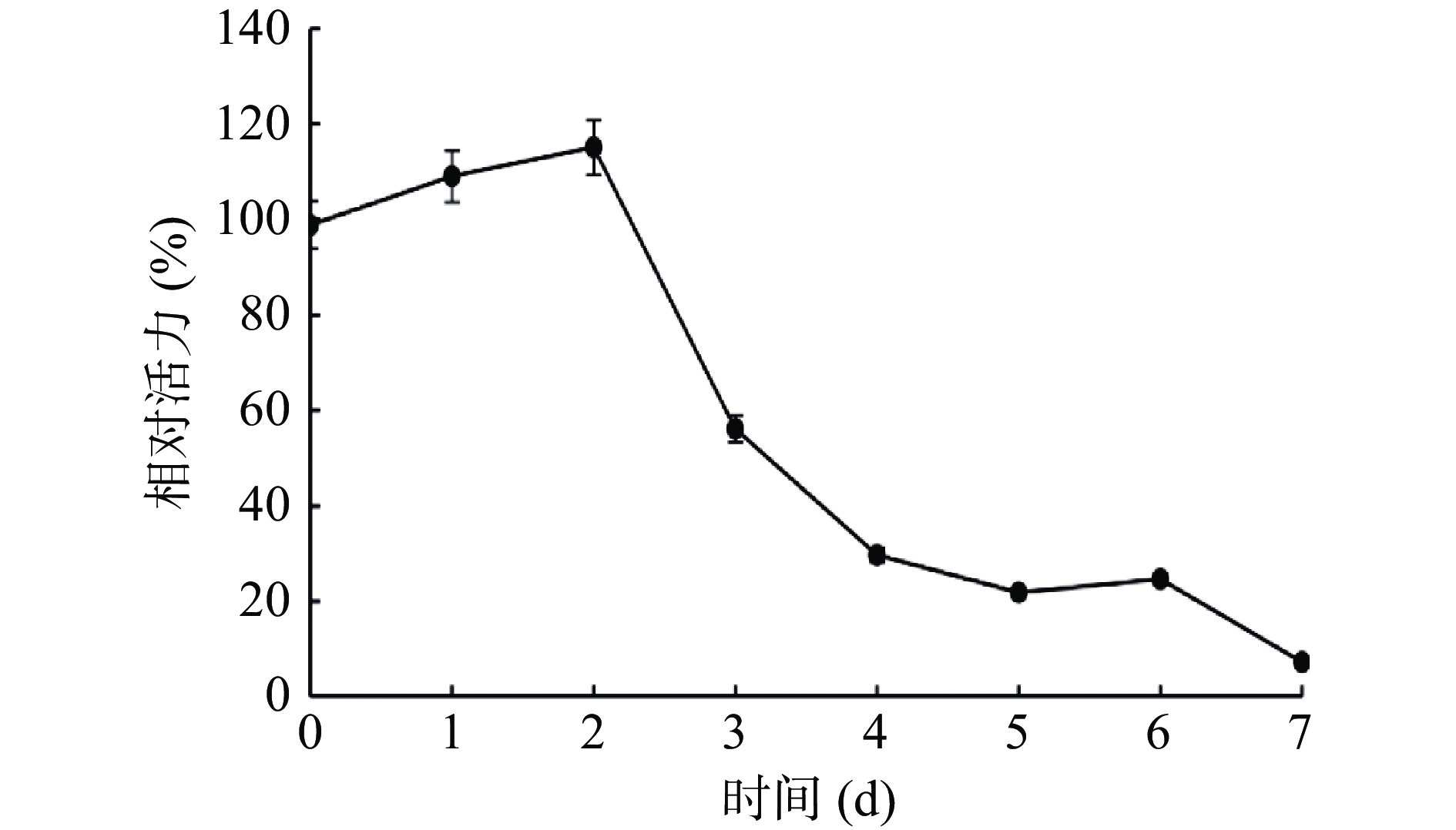
对冷藏过程中虾夷扇贝丝氨酸蛋白酶的酶活力进行测定(图2)。结果发现,丝氨酸蛋白酶酶活力在2 d后开始下降。内源蛋白酶水解蛋白质引起肌肉蛋白结构的破环与质地下降密切关联[10-11]。内源性蛋白酶中,最适pH为弱碱性的丝氨酸蛋白酶可同时降解肌原纤维蛋白和结缔组织蛋白[28]。对海湾扇贝的相关研究结果显示,丝氨酸蛋白酶抑制剂PMSF显著抑制了扇贝冷藏过程中肌球蛋白重链(MHC)和肌动蛋白(Actin)的降解,说明丝氨酸蛋白酶是导致肌原纤维主要结构蛋白降解的酶[16]。因此,内源性丝氨酸蛋白酶值得重点关注。
![]() 图 2 虾夷扇贝肌肉冷藏过程中丝氨酸蛋白酶活力的变化Figure 2. Changes of SP activity in Yesso scallop muscle during cold storage
图 2 虾夷扇贝肌肉冷藏过程中丝氨酸蛋白酶活力的变化Figure 2. Changes of SP activity in Yesso scallop muscle during cold storage2.2 丝氨酸蛋白酶的纯化结果
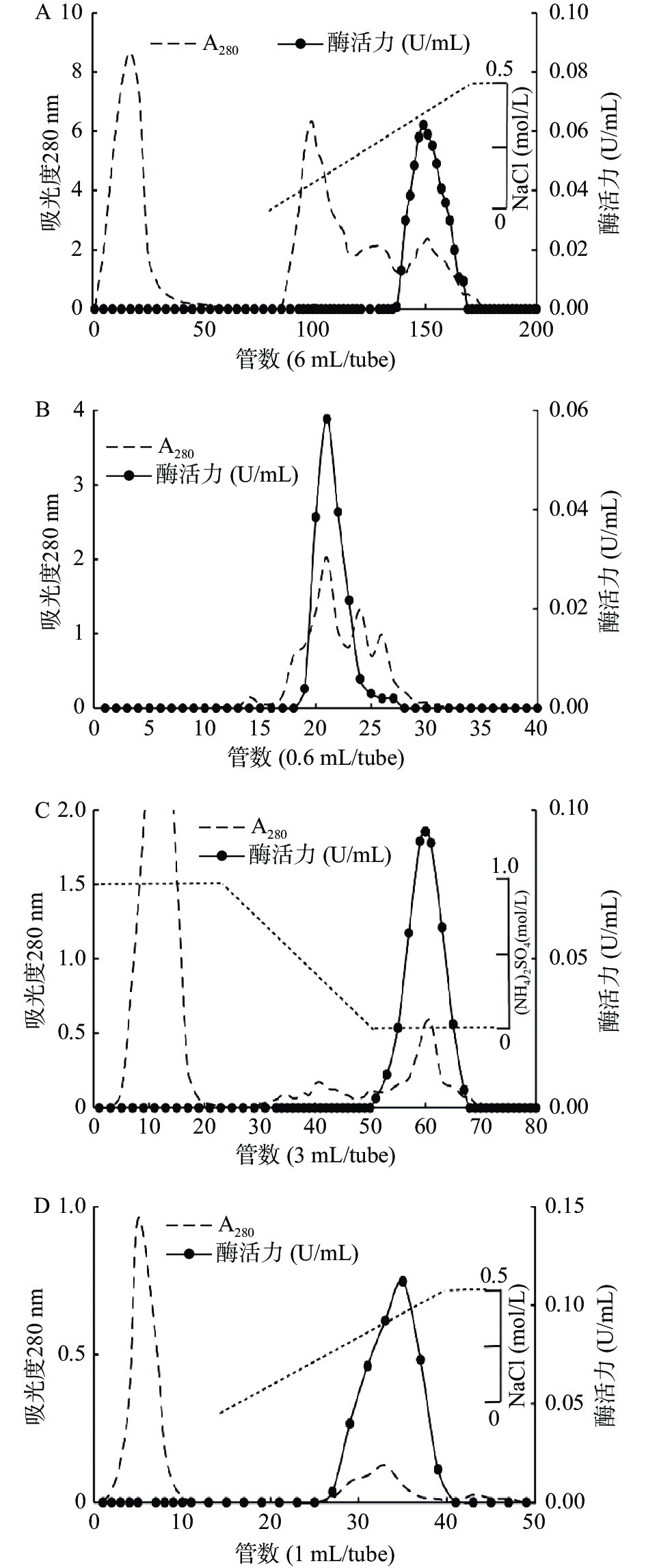
通过30%~60%饱和度硫酸铵盐析从虾夷扇贝肌肉中提取SP粗酶液,上样于预先平衡好的DEAE-Sepharose阴离子交换层析柱。图3A显示,酶活性峰在0.25~0.5 mol/L NaCl下被洗脱下来。将样品浓缩后经Superdex 200凝胶过滤层析柱,可去除小部分的低分子量蛋白(图3B)。收集活性峰后上样于Phenyl-Sepharose疏水层析柱,目的蛋白酶在(NH4)2SO4浓度为0 mol/L时被洗脱下来(图3C)。收集活性峰后上样于DEAE-Sepharose(图3D),收集25~40管样品,用10 kDa超滤浓缩管浓缩后进行SDS-PAGE分析,经过连续柱层析可获得高纯度目的蛋白酶样品用于后续实验。
![]() 图 3 丝氨酸蛋白酶分离纯化柱层析图注:(A) DEAE-Sepharose; (B) Superdex 200; (C) Phenyl-Fast Flow; (D) DEAE-Sepharose。Figure 3. Chromatography for purification of SP
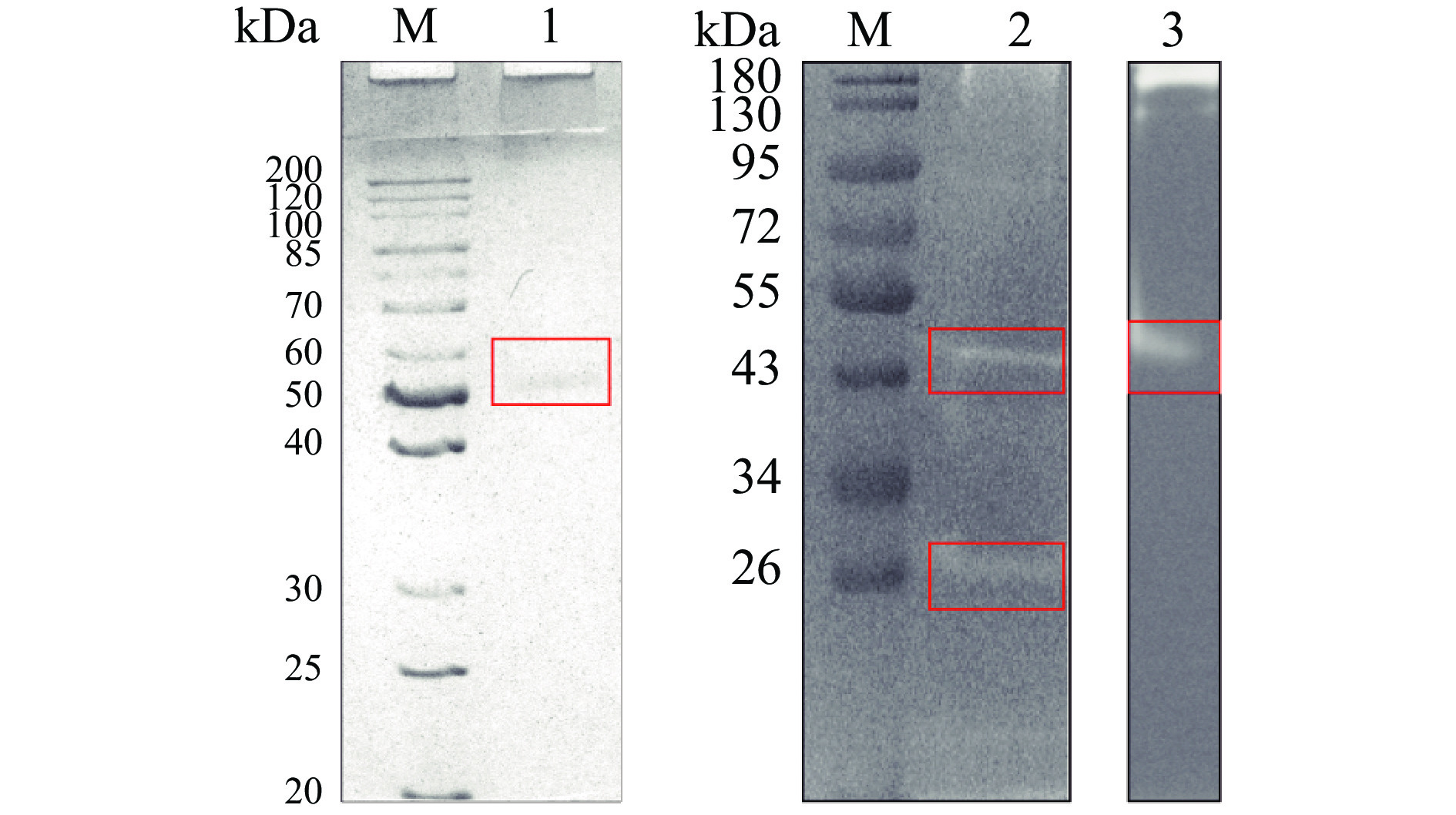
图 3 丝氨酸蛋白酶分离纯化柱层析图注:(A) DEAE-Sepharose; (B) Superdex 200; (C) Phenyl-Fast Flow; (D) DEAE-Sepharose。Figure 3. Chromatography for purification of SP对纯化蛋白的SDS-PAGE和明胶酶谱分析结果显示,该酶主要以分子量为52 kDa的形式存在。但在还原条件下明胶酶谱结果出现有降解明胶的分子量为52 kDa和26 kDa的2个条带(图4,泳道2),推测其在天然条件下存在两种结构,即单体和二聚体,单体的分子量为26 kDa,二聚体的分子量为52 kDa(图4)。其单体分子量26 kDa和南美白对虾(28 kDa)[30]消化腺中分离纯化得到的丝氨酸蛋白酶相近,但与皱纹盘鲍(34 kDa)[31]中克隆表达得到的丝氨酸蛋白酶不同,提示不同物种中丝氨酸蛋白酶的差异性。
![]() 图 4 虾夷扇贝SP的SDS-PAGE和明胶酶谱分析注:M:标准蛋白;1:SP, SDS-PAGE;2:SP+β-Me,明胶酶谱;3:SP-β-Me,明胶酶谱。Figure 4. SDS-PAGE and gelatin zymography of SP from Yesso scallop
图 4 虾夷扇贝SP的SDS-PAGE和明胶酶谱分析注:M:标准蛋白;1:SP, SDS-PAGE;2:SP+β-Me,明胶酶谱;3:SP-β-Me,明胶酶谱。Figure 4. SDS-PAGE and gelatin zymography of SP from Yesso scallop2.3 SP的酶学性质分析
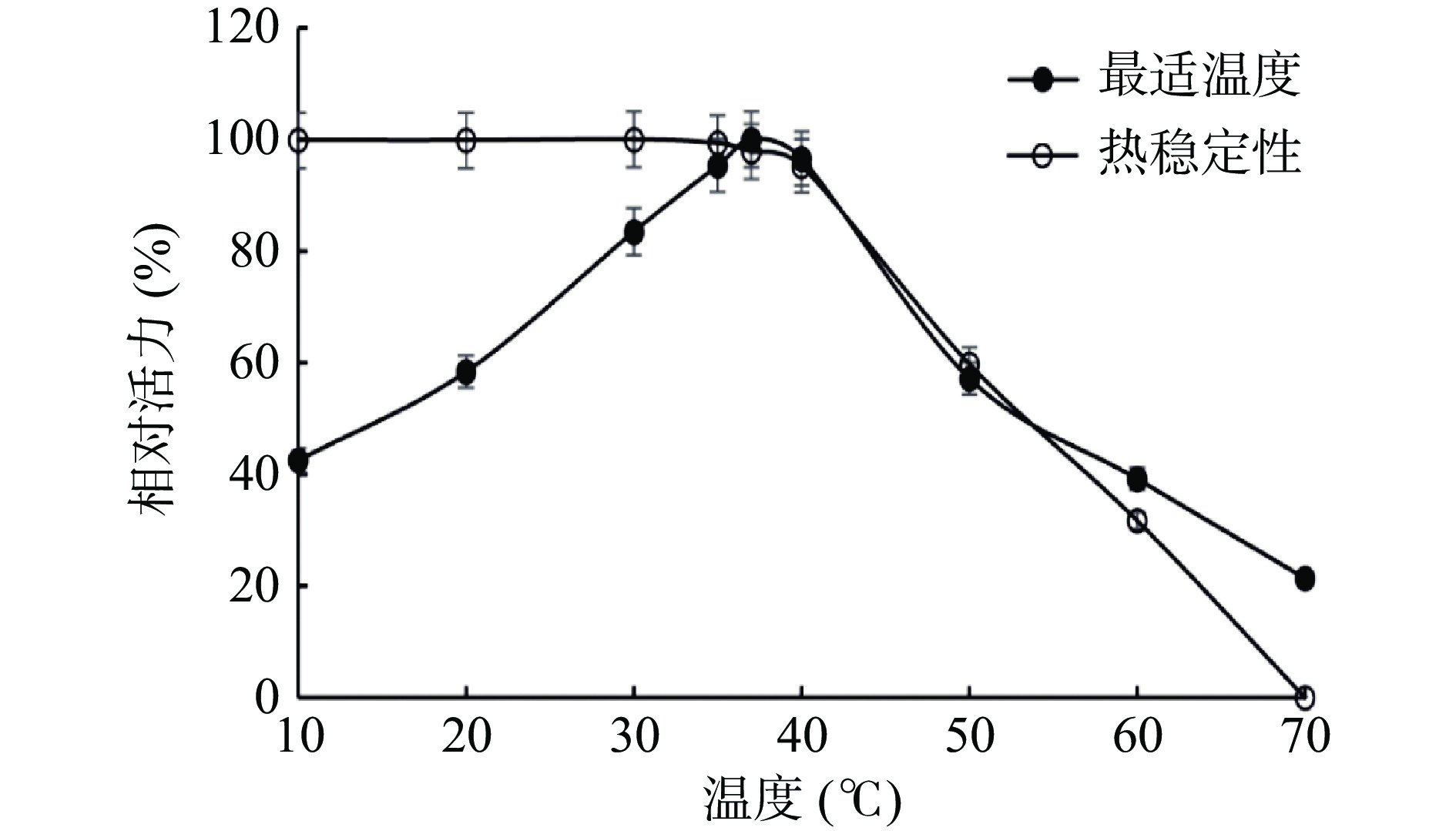
2.3.1 最适温度和热稳定性
由图5可知,SP酶活力的最适温度为37 ℃,即便在10 ℃时,SP仍有最高活性的40%。根据热稳定性结果分析,在温度40 ℃以下,酶活力能够保持较高水平,40 ℃以上酶活力急剧降低,70 ℃时SP基本失活。说明该酶属于低温酶,在低温下能较好保持丝氨酸蛋白酶酶活力。该结果与来自南美白对虾[30]、蓝圆鲹[32]等水产动物中的丝氨酸蛋白酶性质相似,都属于低温酶,应与它们生活环境温度有关。
![]() 图 5 虾夷扇贝丝氨酸蛋白酶的最适温度及热稳定性分析Figure 5. Optimum temperature and thermal stability of SP from Yesso scallop
图 5 虾夷扇贝丝氨酸蛋白酶的最适温度及热稳定性分析Figure 5. Optimum temperature and thermal stability of SP from Yesso scallop2.3.2 SP最适pH和pH稳定性
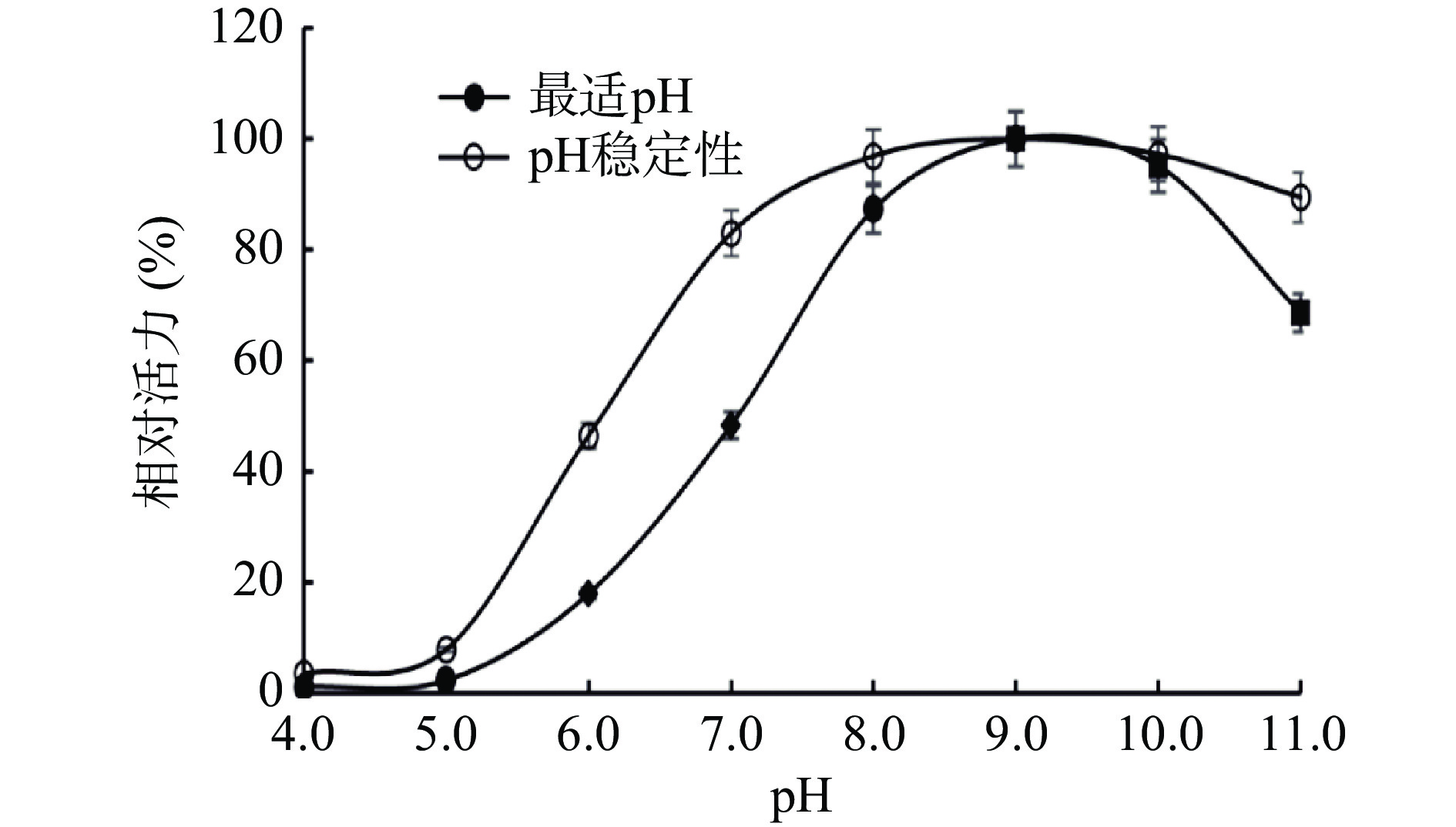
由SP的pH性质分析结果可知,图6表明,SP在pH9.0条件下相对酶活力最高,即pH9.0为SP最适pH;在pH8.0~11.0条件范围内,SP的酶活力都较为稳定,当pH小于7.0时,SP酶活力急剧下降,在pH6.0条件下SP活性只有18%。在pH稳定性方面,SP在pH7.0~11.0条件下,酶活力能够稳定保持在较高水平;pH5.0时,SP活性基本失活。表明SP为一种弱碱性酶。这和南美白对虾[30]消化腺中SP及蓝圆鲹[32]中SP结果相似。但虾夷扇贝SP的耐碱性更好,在pH8.0~11.0范围内,SP酶活力都能稳定保持在80%以上,而南美白对虾[30]SP在pH11.0时,其酶活力基本失活。SP为弱碱性蛋白酶的性质可能对肌肉在冷藏过程前期肌原纤维蛋白降解有影响。
![]() 图 6 虾夷扇贝丝氨酸蛋白酶的最适pH及pH稳定性分析Figure 6. Optimum pH and pH stability of SP from Yesso scallop
图 6 虾夷扇贝丝氨酸蛋白酶的最适pH及pH稳定性分析Figure 6. Optimum pH and pH stability of SP from Yesso scallop2.3.3 底物特异性研究
为了分析SP的底物特异性,选用几种不同类型的荧光底物,结果如表2所示。结果表明,SP特异性水解羧基侧P1位置含有精氨酸或赖氨酸残基的底物,特别是对Boc-Gln-Ala-Arg-MCA的相对活力最高,其次是Boc-Leu-Lys-Arg-MCA、Boc-Ala-Gly-Pro-Arg-MCA和Boc-Phe-Ser-Arg-MCA,对Boc-Val-Pro-Arg-MCA、Boc-Glu-Lys-Lys-MCA等类蛋白酶的水解率分别为82%和36%。而该酶对糜蛋白酶的底物Suc-Leu-Leu-Val-Tyr-MCA或Suc-Ala-Ala-Pro-MCA无活性。该酶也不能水解氨肽酶底物Arg-MCA。与大多数水产动物丝氨酸蛋白酶结果相似,SP对组织蛋白酶类底物(Z-Phe-Arg-MCA、Z-Arg-Arg-MCA)也有微弱的水解能力。底物特异性表明该酶为类胰蛋白酶丝氨酸蛋白酶。
表 2 虾夷扇贝丝氨酸蛋白酶的底物特异性分析Table 2. Substrate characterization of the SP from Yesso scallop底物(10 µmol/L) 相对活力(%) Boc-Phe-Ser-Arg-MCA 100±0.0 Boc-Gln-Ala-Arg-MCA 123±0.3 Boc-Leu-Lys-Arg-MCA 116±0.4 Boc-Ala-Gly-Pro-Arg-MCA 108±0.2 Boc-Val-Pro-Arg-MCA 82±1.2 Boc-Glu-Lys-Lys-MCA 36±0.4 Z-Phe-Arg-MCA 36±2.3 Z-Arg-Arg-MCA 37±0.9 Suc-Leu-Leu-Val-Tyr-MCA 0 Suc-Ala-Ala-Pro-MCA 0 Arg-MCA 0 2.3.4 不同蛋白酶抑制剂对SP的影响
由表3可知,丝氨酸蛋白酶抑制剂Pefabloc SC、Benzamidine和胰蛋白酶抑制剂Leupeptin均能明显抑制SP活性。当PMSF浓度为1 mmol/L时,对SP的抑制作用不明显,但当浓度增至10 mmol/L时,几乎能够完全抑制其活性。金属离子蛋白酶抑制剂EGTA和1,10-phenathroline对丝氨酸蛋白酶均有不同程度的抑制作用。半胱氨酸蛋白酶抑制剂E-64对该酶活力有微弱的抑制作用。结合底物特异性和蛋白酶抑制剂结果表明,SP为一种类胰蛋白酶的丝氨酸蛋白酶。
表 3 蛋白酶抑制剂对丝氨酸蛋白酶活力的影响Table 3. Inhibitory effect of different proteinase inhibitors on the activity of SP抑制剂 浓度(mmol/L) 相对活力(%) control 0 100±0.0 EGTA 1 47±0.2 EDTA 1 90±0.1 10 93±0.3 1,10-phenanthroline 1 96±0.2 5 84±0.2 PMSF 1 86±0.1 10 3±0.4 E-64 0.015 78±0.1 0.1 80±0.1 Benzazmidine 0.5 15±0.3 5 7±0.2 Pefabloc SC 0.2 11±0.5 2 3±0.1 Leupeptin 0.01 3±0.1 0.03 2±0.6 2.3.5 不同金属离子对SP的影响
由抑制剂结果得知SP酶活力可能与金属离子有关,由表4可知,Fe2+、Zn2+、Cu2+能够明显抑制SP的酶活力,在0.1 mmol/L时,Fe2+可抑制其初始活性的83%。Zn2+对SP酶活力的抑制作用与浓度相关,0.01 mmol/L时只抑制其活性10%,0.05 mmol/L时抑制其64%活性,浓度增至0.1 mmol/L抑制84%的SP活性。0.01 mmol/L Ca2+能够抑制SP约50%的酶活力;0.1 mmol/L的Ba2+能抑制约20%酶活;而Mg2+、Mn2+对SP酶活力几乎没有明显的抑制作用。该结果与蓝圆鲹中SP的性质有差异,表明不同物种SP其性质略有不同[32]。
表 4 金属离子对丝氨酸蛋白酶活力的影响Table 4. Effect of metal ions on the activity of SP金属离子 浓度(mmol/L) 相对活力(%) control 0 100±0.0 Ca2+ 0.01 58±0.4 0.05 72±0.1 0.1 75±0.2 Fe2+ 0.01 78±0.3 0.05 73±0.5 0.1 17±0.4 Zn2+ 0.01 90±0.1 0.05 36±0.3 0.1 16±0.3 Mg2+ 0.01 96±0.3 0.05 95±0.1 0.1 91±0.2 Ba2+ 0.01 98±0.5 0.05 89±0.2 0.1 81±0.2 Cu2+ 0.01 55±0.3 0.05 72±0.1 0.1 11±0.5 Mn2+ 0.01 95±0.3 0.05 97±0.5 0.1 96±0.5 2.3.6 丝氨酸蛋白酶对虾夷扇贝肌肉肌原纤维的分解
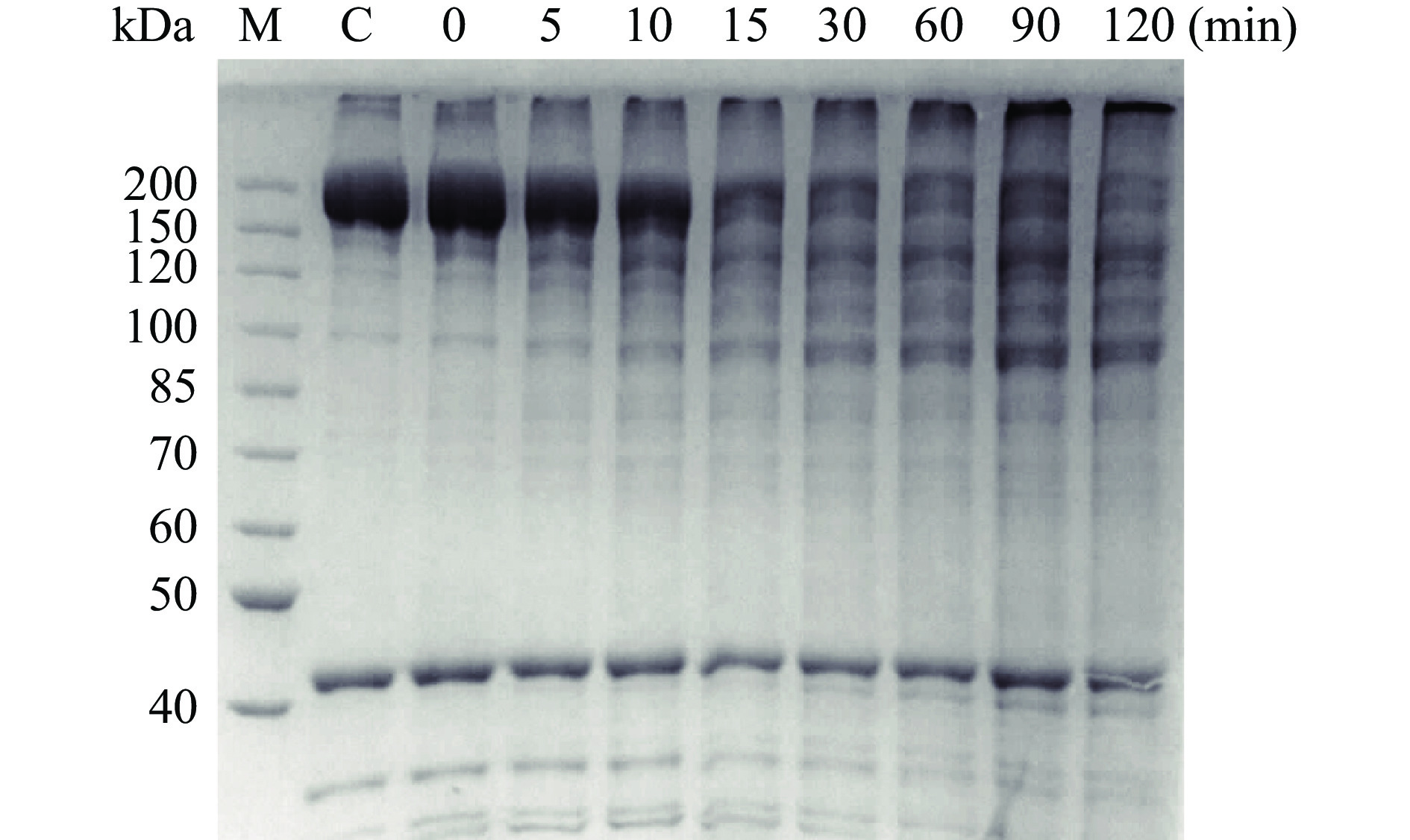
为探究丝氨酸蛋白酶对虾夷扇贝肌原纤维蛋白的作用,将扇贝肌原纤维蛋白后与丝氨酸蛋白酶在37 ℃下孵育后进行SDS-PAGE分析。由图7可知,随着反应时间延长,肌原纤维蛋白逐渐被降解为小分子蛋白片段。反应120 min后,肌球蛋白重链几乎被完全降解。结果证明,丝氨酸蛋白酶是参与虾夷扇贝肌肉蛋白降解的主要蛋白酶之一,对虾夷扇贝肌肉冷藏过程中质构变化有显著的影响。
![]() 图 7 丝氨酸蛋白酶对虾夷扇贝肌肉肌原纤维的分解作用Figure 7. Effect of SP on myofibrillar proteins of Yesso scallop muscle
图 7 丝氨酸蛋白酶对虾夷扇贝肌肉肌原纤维的分解作用Figure 7. Effect of SP on myofibrillar proteins of Yesso scallop muscle3. 结论
虾夷扇贝肌肉在4 ℃的冷藏过程中,质地逐渐劣化,肌肉结构蛋白逐渐降解,特别是肌球蛋白重链有明显的降解趋势,丝氨酸蛋白酶活力整体呈持续下降趋势。从虾夷扇贝肌肉中分离纯化得到一种丝氨酸蛋白酶,推测其单体的分子量为26 kDa,二聚体的分子量为52 kDa。SP的最适温度为37 ℃,最适pH为9.0,特异性水解羧基侧P1位含有精氨酸或赖氨酸残基的底物。丝氨酸蛋白酶特异性抑制剂(Pefabloc SC、Benzamidine、PMSF)和胰蛋白酶抑制剂(Leupeptin)能明显抑制SP的酶活力;金属离子Fe2+、Zn2+、Cu2+对SP有明显的抑制作用。从SP对虾夷扇贝肌肉的分解作用可推测其参与扇贝死后肌肉结构蛋白自身降解,进而影响其肌肉质构。但关于SP的更详细情况,如一级结构,内源性抑制剂存在情况等需要进一步研究。
-
![]()
图 1 虾夷扇贝冷藏过程中肌肉蛋白的自身降解(A)和光密度分析(B)
注:M:标准蛋白。
Figure 1. Protein degradation in Yesso scallop muscle during cold storage (A) and densitometry analysis (B)
![]()
图 2 虾夷扇贝肌肉冷藏过程中丝氨酸蛋白酶活力的变化
Figure 2. Changes of SP activity in Yesso scallop muscle during cold storage
![]()
图 3 丝氨酸蛋白酶分离纯化柱层析图
注:(A) DEAE-Sepharose; (B) Superdex 200; (C) Phenyl-Fast Flow; (D) DEAE-Sepharose。
Figure 3. Chromatography for purification of SP
![]()
图 4 虾夷扇贝SP的SDS-PAGE和明胶酶谱分析
注:M:标准蛋白;1:SP, SDS-PAGE;2:SP+β-Me,明胶酶谱;3:SP-β-Me,明胶酶谱。
Figure 4. SDS-PAGE and gelatin zymography of SP from Yesso scallop
![]()
图 5 虾夷扇贝丝氨酸蛋白酶的最适温度及热稳定性分析
Figure 5. Optimum temperature and thermal stability of SP from Yesso scallop
![]()
图 6 虾夷扇贝丝氨酸蛋白酶的最适pH及pH稳定性分析
Figure 6. Optimum pH and pH stability of SP from Yesso scallop
![]()
图 7 丝氨酸蛋白酶对虾夷扇贝肌肉肌原纤维的分解作用
Figure 7. Effect of SP on myofibrillar proteins of Yesso scallop muscle
表 1 冷藏过程中虾夷扇贝肌肉质构参数变化
Table 1 The change of texture parameters in Yesso callop muscle during cold storage
时间 (d) 硬度 (gf) 咀嚼性(gf) 粘性(gf) 弹性(gf) 0 670±42b 255±18 a 321±2 b 0.34±0.07b 1 736±52a 176±11b 372±17a 0.37±0.04a 2 584±45c 152±8c 276±19c 0.33±0.02b 3 457±38d 132±4d 253±11d 0.32±0.03b 4 330±33e 126±5e 227±12e 0.31±0.02c 5 206±25f 115±2f 176±15f 0.28±0.01d 6 146±27g 102±3g 107±8g 0.27±0.03e 7 68±12h 93±4g 99±7g 0.25±0.02f 注:同列不同小写字母表示差异显著(P<0.05)。  下载: 导出CSV
下载: 导出CSV
表 2 虾夷扇贝丝氨酸蛋白酶的底物特异性分析
Table 2 Substrate characterization of the SP from Yesso scallop
底物(10 µmol/L) 相对活力(%) Boc-Phe-Ser-Arg-MCA 100±0.0 Boc-Gln-Ala-Arg-MCA 123±0.3 Boc-Leu-Lys-Arg-MCA 116±0.4 Boc-Ala-Gly-Pro-Arg-MCA 108±0.2 Boc-Val-Pro-Arg-MCA 82±1.2 Boc-Glu-Lys-Lys-MCA 36±0.4 Z-Phe-Arg-MCA 36±2.3 Z-Arg-Arg-MCA 37±0.9 Suc-Leu-Leu-Val-Tyr-MCA 0 Suc-Ala-Ala-Pro-MCA 0 Arg-MCA 0
下载: 导出CSV
表 3 蛋白酶抑制剂对丝氨酸蛋白酶活力的影响
Table 3 Inhibitory effect of different proteinase inhibitors on the activity of SP
抑制剂 浓度(mmol/L) 相对活力(%) control 0 100±0.0 EGTA 1 47±0.2 EDTA 1 90±0.1 10 93±0.3 1,10-phenanthroline 1 96±0.2 5 84±0.2 PMSF 1 86±0.1 10 3±0.4 E-64 0.015 78±0.1 0.1 80±0.1 Benzazmidine 0.5 15±0.3 5 7±0.2 Pefabloc SC 0.2 11±0.5 2 3±0.1 Leupeptin 0.01 3±0.1 0.03 2±0.6
下载: 导出CSV
表 4 金属离子对丝氨酸蛋白酶活力的影响
Table 4 Effect of metal ions on the activity of SP
金属离子 浓度(mmol/L) 相对活力(%) control 0 100±0.0 Ca2+ 0.01 58±0.4 0.05 72±0.1 0.1 75±0.2 Fe2+ 0.01 78±0.3 0.05 73±0.5 0.1 17±0.4 Zn2+ 0.01 90±0.1 0.05 36±0.3 0.1 16±0.3 Mg2+ 0.01 96±0.3 0.05 95±0.1 0.1 91±0.2 Ba2+ 0.01 98±0.5 0.05 89±0.2 0.1 81±0.2 Cu2+ 0.01 55±0.3 0.05 72±0.1 0.1 11±0.5 Mn2+ 0.01 95±0.3 0.05 97±0.5 0.1 96±0.5
下载: 导出CSV
-
[1] 王丹, 吴反修. 中国渔业统计年鉴[G]. 北京: 中国农业出版社, 2021 WANG Dan, WU Fanxiu. China fishery statistics yearbook[G]. Beijing: China Agriculture Press, 2021.
[2] 汤俊杰, 段振华, 杨丽娟. 扇贝的加工利用研究进展[J]. 食品工业,2013,34(5):191−194. [TANG Junjie, DUAN Zhenhua, YANG Lijuan. Research progress on processing and utilization of scallop[J]. Food Industry,2013,34(5):191−194. TANG Junjie, DUAN Zhenhua, YANG Lijuan. Research progress on processing and utilization of scallop[J]. Food Industry, 2013, 34(5): 191-194.
[3] 高加龙, 章超桦, 秦小明, 等. 虾夷扇贝内脏团对镉的富集特性及生理响应[J]. 广东海洋大学学报,2017,32(3):204−210. [GAO Jialong, ZHANG Chaohua, QIN Xiaoming, et al. Cadmium enrichment characteristics and physiological response of visceral mass of scallop prawn[J]. Journal of Guangdong Ocean University,2017,32(3):204−210. GAO Jialong, ZHANG Chaohua, QIN Xiaoming, et al. Cadmium enrichment characteristics and physiological response of visceral mass of scallop prawn[J]. Journal of Guangdong Ocean University, 2017, 32(3): 204-210.
[4] 刘征. 扇贝干燥关键技术研究[D]. 保定: 河北农业大学, 2012 LIU Zheng. Study on key drying technology of scallop[D]. Baoding: Agricultural University of Hebei, 2012.
[5] 陈文秀. 虾夷扇贝保活品质变化与贮藏条件调控技术研究[D]. 上海: 上海海洋大学, 2020 CHEN Wenxiu. Study on quality change and storage condition control technology of scallop shrimp[D]. Shanghai: Shanghai Ocean University, 2020.
[6] 姜明慧, 田元勇, 闫丽新, 等. 冷藏过程中虾夷扇贝横纹肌微观结构变化与肌原纤维蛋白稳定性[J]. 食品科学,2020,41(21):175−181. [JIANG Minghui, TIAN Yuanyong, YAN Lixin, et al. Microstructural changes of striated muscle and stability of myofibrin in scallop shrimp during cold storage[J]. Food Science,2020,41(21):175−181. doi: 10.7506/spkx1002-6630-20191016-159 JIANG Minghui, TIAN Yuanyong, YAN Lixin, et al. Microstructural changes of striated muscle and stability of myofibrin in scallop shrimp during cold storage[J]. Food Science, 2020, 41(21): 175-181. doi: 10.7506/spkx1002-6630-20191016-159
[7] 徐锦华, 孟泽玲, 葛诗琪, 等. 刺参体壁胰/类胰丝氨酸蛋白酶的性质及其在自溶中的作用[J]. 食品科学,2019,40(18):95−100. [XU Jinhua, MENG Zeling, GE Shiqi, et al. Characterization of trypsin/trypsine-like protease in body wall of Stichopus japonicus and its role in autolysis[J]. Food Science,2019,40(18):95−100. doi: 10.7506/spkx1002-6630-20181004-019 XU Jinhua, MENG Zeling, GE Shiqi, et al. Characterization of trypsin/trypsine-like protease in body wall of Stichopus japonicus and its role in autolysis[J]. Food Science, 2019, 40(18): 95-100. doi: 10.7506/spkx1002-6630-20181004-019
[8] CHENG J H, SUN D W, HAN Z, et al. Texture and structure measurements and analyses for evaluation of fish and fillet freshness quality: A review[J]. Comprehensive Reviews in Food Science and Food Safety,2014,13(1):52−61. doi: 10.1111/1541-4337.12043
[9] TANIOKA T, HATTORI A, MIZUTANI S. Regulation of the human leukocyte-derived argine aminopeptidase/endoplasmic reticulum-aminopeptidase 2 gene by interferon-C[J]. FEBS Journal,2005,272:916−928. doi: 10.1111/j.1742-4658.2004.04521.x
[10] SINGH A, BENJAKUL S. Proteolysis and its control using protease inhibitors in fish and fish products: A review[J]. Comprehensive Reviews in Food Science and Food Safety,2018,17(2):496−509. doi: 10.1111/1541-4337.12337
[11] YANG F, RUSTAD T, XU Y, et al. Endogenous proteolytic enzymes-a study of their impact on cod (Gadus morhua) muscle proteins and textural properties in a fermented product[J]. Food Chemistry,2015,172:551−558. doi: 10.1016/j.foodchem.2014.09.086
[12] BROWN M R, SIKES A L, ELLIOTT N G, et al. Physicochemical factors of abalone quality: A review[J]. Journal of Shellfish Research,2008,27(4):835−842. doi: 10.2983/0730-8000(2008)27[835:PFOAQA]2.0.CO;2
[13] SRIKET C, BENJAKUL S, VISESSANGUAN W, et al. Collagenolytic serine protease in fresh water prawn (Macrobrachium rosenbergii): Characteristics and its impact on muscle during iced storage[J]. Food chemistry,2011,124(1):29−35. doi: 10.1016/j.foodchem.2010.05.098
[14] LIU J, YOSHIDA A, GAO Y, et al. Purification and characterization of a sarcoplasmic serine proteinase from threadfin bream nemipterus virgatus muscle[J]. Food Chemistry,2019,284:198−204. doi: 10.1016/j.foodchem.2019.01.024
[15] 杜翠红, 曹敏杰. 鱼类肌原纤维结合型丝氨酸蛋白酶研究进展[J]. 食品科学,2013,34(9):336−339. [DU Cuihong, CAO Minjie. Research progress of fish myofibrillary binding serine protease[J]. Food Science,2013,34(9):336−339. doi: 10.7506/spkx1002-6630-201309067 DU Cuihong, CAO Minjie. Research progress of fish myofibrillary binding serine protease[J]. Food Science, 2013, 34(9): 336-339. doi: 10.7506/spkx1002-6630-201309067
[16] LIU B, LIU Z Q, LI D Y, et al. Action of endogenous proteases on texture deterioration of the bay scallop (Argopecten irradians) adductor muscle during cold storage and its mechanism[J]. Food Chemistry,2020,323:126790. doi: 10.1016/j.foodchem.2020.126790
[17] KWON T, KIM M, CHOI H, et al. A masquerade-like serine protease homolog is necessary for prophenoloxidase activity in the coleopteran insect, holotrichia diomphalia larvae[J]. European Journal of Biochemistry,2000,267:6188−6196. doi: 10.1046/j.1432-1327.2000.01695.x
[18] DELOTTO Y, DELOTTO R. Proteolytic processing of the drosophila spatzle protein by easter generates a dimeric ngf-like molecule with ventralising activity[J]. Mechanisms of Development,1998,72(1-2):141−148. doi: 10.1016/S0925-4773(98)00024-0
[19] KAWABATA S, TOKUNAGA F, KUGI Y, et al. Limulus factor D, a 43-kDa protein isolated from horseshoe crab hemocytes, is a serine protease homologue with antimicrobial activity[J]. FEBS Letters,1996,398(2-3):146−150. doi: 10.1016/S0014-5793(96)01224-0
[20] 游银川. 皱纹盘鲍肌肉质构相关蛋白酶的研究[D]. 厦门: 集美大学, 2016 YOU Yinchuan. Study on proteinases related to texture from the muscle of abalone (Haliotis discus hannai)[D]. Xiamen: Jimei University, 2016.
[21] 林怡晨, 刘伟峰, 孙小舒, 等. 蓝圆鲹分离蛋白水溶性蛋白酶的鉴定及性质[J]. 集美大学学报(自然科学版),2020,25(2):97−104. [LIN Yichen, LIU Weifeng, SUN Xiaoshu, et al. Identification and characterization of water-soluble protease in scad[J]. Journal of Jimei University (Natural Science Edition),2020,25(2):97−104. LIN Yichen, LIU Weifeng, SUN Xiaoshu, et al. Identification and characterization of water-soluble protease in scad[J]. Journal of Jimei University (Natural Science Edition), 2020, 25(2): 97-104.
[22] 麻金花, 翁凌, 颜龙杰, 等. 海参内脏胶原降解酶的纯化及性质研究[J]. 水产学报,2015,39(7):34−37. [MA Jinhua, WENG Ling, YAN Longjie, et al. Purification and characterization of collagen degrading enzymes from sea cucumber viscera[J]. Journal of Fisheries of China,2015,39(7):34−37. MA Jinhua, WENG Ling, YAN Longjie, et al. Purification and characterization of collagen degrading enzymes from sea cucumber viscera[J]. Journal of Fisheries of China, 2015, 39(7): 34-37.
[23] CAO M J, SHAO W, LI Y, et al. Identification of a myofibril-bound serine proteinase in the skeletal muscle of silver carp[J]. Journal of Food Biochemistry,2004,28(5):373−386. doi: 10.1111/j.1745-4514.2004.04203.x
[24] XIONG X, HE B, JIANG D, et al. Postmortem biochemical and textural changes in the patinopecten yessoensis adductor muscle (Pyam) during iced storage[J]. International Journal of Food Properties,2019,22(1):1024−1034. doi: 10.1080/10942912.2019.1625367
[25] AYALA M D, ABDEL I, SANTAELLA M, et al. Muscle tissue structural changes and texture development in sea bream, Sparus aurata L., during post-mortem storage[J]. LWT-Food Science and Technology,2010,43(3):465−475. doi: 10.1016/j.lwt.2009.08.023
[26] SHARIFIMEHR S, SOLTANIZADEH N, HOSSEIN GOLI S A. Effects of edible coating containing nano-emulsion of aloe vera and eugenol on the physicochemical properties of shrimp during cold storage[J]. Journal of the Science of Food and Agriculture,2019,99(7):3604−3615. doi: 10.1002/jsfa.9581
[27] HULTMANN L, RUSTAD T. Textural changes during iced storage of salmon (Salmo salar) and cod (Gadus morhua)[J]. Journal of Aquatic Food Product Technology,2002,11(3-4):105−123. doi: 10.1300/J030v11n03_09
[28] TAKAHASHI K, KUROSE K, OKAZAKI E, et al. Effect of various protease inhibitors on heat-induced myofibrillar protein degradation and gel-forming ability of red tilefish (Branchiostegus japonicus) meat[J]. LWT-Food Science and Technology,2016,68:717−723. doi: 10.1016/j.lwt.2016.01.022
[29] LIU B, LIU Y X, LIU Z Q, et al. Effects of natural trypsin inhibitor fromssoybean on texture deterioration of the bay scallop (Argopecten irradians) during cold storage and its mechanism[J]. International Journal of Food Science & Technology,2020,55(11):3432−3440.
[30] 翁凌, 李腾, 阴利华, 等. 南美白对虾丝氨酸蛋白酶的分离纯化及性质研究[J]. 集美大学学报(自然科学版),2010,15(4):272−278. [WENG Ling, LI Teng, YIN Lihua, et al. Isolation, purification and characterization of serine protease from Penaeus vannamei[J]. Journal of Jimei University (Natural Science Edition),2010,15(4):272−278. WENG Ling, LI Teng, YIN Lihua, et al. Isolation, purification and characterization of serine protease from Penaeus vannamei[J]. Journal of Jimei University (Natural Science Edition), 2010, 15(4): 272-278.
[31] 胡健健. 皱纹盘鲍丝氨酸蛋白酶的基因克隆、体外表达及其免疫调控作用[D]. 厦门: 集美大学, 2017 HU Jianjian. Gene cloning, in vitro expression and immune regulation of abalone serine protease[D]. Xiamen: Jimei University, 2017.
[32] 王梦想, 钟婵, 蔡秋凤, 等. 蓝圆肌肉中丝氨酸蛋白酶的分离纯化及性质研究[J]. 食品工业科技,2012,29(19):58−67. [WANG Mengxiang, ZHONG Chan, CAI Qiufeng, et al. Purification and characterization of serine protease from blue circle muscle[J]. Science and Technology of food Industry,2012,29(19):58−67. WANG Mengxiang, ZHONG Chan, CAI Qiufeng, et al. Purification and characterization of serine protease from blue circle muscle[J]. Science and Technology of food Industry, 2012, 29(19): 58-67.
 下载:
下载:







计量
- 文章访问数:
- HTML全文浏览量:
- PDF下载量:
