


The Effect of Polysaccharide of Poria cocos on Key Metabolites of Bifidobacterium BB-12
-
摘要: 为探究动物双岐杆菌乳双歧亚种BB-12代谢茯苓多糖的关键代谢产物,采用基于液质联用技术(LC-MS/MS)的非靶向代谢组学手段,以葡萄糖作为对照,分析茯苓多糖对动物双岐杆菌乳双歧亚种BB-12关键代谢产物的影响。结果表明,茯苓多糖可显著促进双歧杆菌BB-12的生长。结合主成分分析(Principal Component Analysis, PCA)和正交偏最小二乘判别模型(Orthogonal Partial Least Squares Discriminant Analysis, OPLS-DA)等分析手段,筛选出丹芝酸、黄藤素等具有明确生理功能的差异代谢产物共12种,并显著富集到ABC转运蛋白通路、亚油酸代谢通路、半乳糖代谢通路等17条差异代谢通路。这些代谢物和代谢通路均可证明双歧杆菌BB-12在发酵茯苓多糖的情况下表现出更优异的生长性能和益生功效。该实验从小分子代谢物水平分析了动物双歧杆菌乳双歧亚种BB-12代谢茯苓多糖后产生的代谢产物,为进一步研究茯苓及动物双岐杆菌乳双歧亚种BB-12在人体内发挥益生功效的相关机制提供理论依据和指导。Abstract: In order to explore the key metabolites of Bifidobacterium animalis spp. lactis BB-12 metabolizing polysaccharides of Poria cocos, a non-targeted metabolomics method based on liquid chromatography-mass spectrometry (LC-MS/MS) was used, and glucose was used as a control. The effects of polysaccharides of Poria cocos on key metabolites of Bifidobacterium animalis spp. lactis BB-12 were analyzed. The results showed that polysaccharides of Poria cocos could significantly promote the growth of Bifidobacterium BB-12. By using component analysis (PCA) and orthogonal partial least squares discriminant analysis (OPLS-DA), some representative differential metabolites were screened out such as palmatine and ganolucidic acid, etc. There were 12 kinds of key differential metabolites, and 17 differential metabolic pathways were significantly enriched such as ABC transporters, linoleic acid metabolism, and galactose metabolism. These metabolites and pathways demonstrated that Bifidobacterium BB-12 exhibited better growth performance and prebiotic efficacy in the case of fermenting polysaccharides of Poria cocos. This experiment analyzed the metabolites produced by polysaccharides of Poria cocos from the level of small molecular metabolites which were metabolized by Bifidobacterium animalis spp. lactis BB-12. The results provide the theoretical basis and guidance for further study on the probiotic effect of polysaccharides of Poria cocos and Bifidobacterium animalis spp. lactis BB-12 in human bodies.
-
茯苓(Poria cocos)是一种我国传统的中药材,也是重要的药食同源功能性食材,具有益脾和胃、宁心安神的功效。它来自于多孔菌科茯苓属真菌茯苓的干燥菌核,多糖为茯苓的主要化学成分之一[1],具有抗炎、抗癌、调节免疫、降血糖等功效[2]。KHAN等[3]对小鼠的肠道菌群分析发现,饲喂茯苓的小鼠肠道内可产生乳酸、短链脂肪酸的菌群以及具有分解多糖、改善肥胖能力的菌群。GAO等[4]证明了茯苓多糖作为一种益生元可以促进双歧杆菌的增殖并产生了大量以乙酸为主的短链脂肪酸。JIANG等[5]的研究表明,茯苓提取物可以在动物体内促进双歧杆菌生长,并抑制沙门氏菌和金黄色葡萄球菌等有害菌的生长。此外,还有一些实验结果显示[6],高剂量的茯苓能显著提高小鼠肠道内双歧杆菌的水平,并具有良好的肠道菌群调节效果,且其药效和剂量有关,提示茯苓有可能作为微生态调节剂或添加剂,用于肠道菌群调节和相关预防治疗。
动物双歧杆菌乳双歧亚种(Bifidobacterium animalis spp. lactis BB-12,下文简称双歧杆菌BB-12)是迄今为止世界上临床研究最为充分的双歧杆菌菌株[7]。研究证明,双歧杆菌BB-12能够改善肠道功能[8-9]、防止腹泻[10]、并能减少抗生素治疗带来的副作用[11]。在免疫功能方面,临床研究表明双歧杆菌BB-12会提高机体对常见呼吸道感染的抵抗力,同时降低急性呼吸道感染的发病率。双歧杆菌BB-12还可通过其代谢物来防止肠道上皮屏障功能受到破坏[12]。此外,双歧杆菌BB-12还有着优良的抑菌性。研究表明,双歧杆菌BB-12对蜡状芽孢杆菌、艰难梭菌、产气荚膜杆菌、大肠杆菌和粪肠球菌等表现出明显的抑菌作用[13]。目前,双歧杆菌BB-12已在食品、保健等多个领域广泛应用。
现有文献中大多将茯苓多糖通过动物实验或临床数据证实茯苓多糖具有重要的生物活性,通过促进肠道有益菌(双歧杆菌等)增殖,进而对疾病进行调控。由于肠道菌群宏基因组测序技术的局限性,目前的体内实验结果仅能反映出体内双歧杆菌等益生菌的丰度和比例上调,很难从益生菌种的水平或株的水平来证明茯苓多糖对其有明显的促进作用,并且这些研究结果具体影响的是哪些种类的双歧杆菌以及产生的具体代谢产物、与免疫等调控相关的代谢途径等机理研究尚未见报道。由此,本研究以茯苓多糖为例,基于非靶向代谢组学技术探究动物双岐杆菌乳双歧亚种BB-12发酵茯苓多糖和葡萄糖代谢产物的差异。拟通过小分子代谢水平解释茯苓多糖在体内分解产生的关键产物及其发挥的生理活性,探究双歧杆菌与益生元复配的条件下发挥益生功效的关键产物及相关潜在机理,为日后深入研究茯苓多糖的生理活性及双歧杆菌的益生机制提供新的科学思路和研究模式。
1. 材料与方法
1.1 材料与仪器
动物双歧杆菌乳双歧亚种BB-12 由东北农业大学乳成分功能与营养实验室保存;MRS肉汤培养基 青岛海博生物技术有限公司;无糖MRS培养基 上海瑞永生物科技有限公司;茯苓多糖(50%) 西安圣青生物科技有限公司;甲醇、乙腈、甲酸 HPLC级,赛默飞世尔科技公司。
UHPLC液相色谱系统、Q-Exactive HF-X质谱仪 赛默飞世尔科技公司;Wonbio-96c多样品冷冻研磨仪 上海万柏生物科技有限公司;Centrifuge 5430 R高速冷冻离心机 德国Eppendorf公司;JXDC-20氮气吹扫仪 上海净信实业发展有限公司;SBL-10 TD控温超声波清洗机 宁波新芝生物科技股份有限公司;Anaero Pack TM-Anaero厌氧产气袋 日本三菱瓦斯化学株式会社。
1.2 实验方法
1.2.1 发酵液的制备
向无糖MRS培养基中加入2%的茯苓多糖作为双歧杆菌BB-12发酵的唯一碳源,然后将MRS肉汤培养基、加入2%茯苓多糖的无糖MRS培养基分别装入5 mL洁净试管中,121 ℃高压灭菌15 min备用。
1.2.2 生长曲线的测定与拟合
−80 ℃保存的动物双歧杆菌乳双歧亚种BB-12使用前均按照2%的比例接种于MRS肉汤培养基中,37 ℃厌氧培养24 h,经2次传代后,取对数生长期菌液分别接种至普通MRS肉汤培养基和加入2%茯苓多糖的无糖MRS培养基中,利用第三代菌种的发酵液进行实验。菌种在37 ℃下恒温厌氧发酵48 h,每4 h在波长600 nm处测定一次发酵液的吸光度及pH,每个样品做4个重复。
常用的微生物生长动力学模型有:Gompertz模型、Logistic模型、Richards模型等[14]。本实验采用Origin Pro 2019 b软件中的SGompertz模型,迭代算法为Levernberg-Marquardt优化算法,表达式如下:
y=A×e(−e(−k×(x−xc))) 式中,A、xc、k为参数[15],A为模型所预测的生长最大值;xc为拐点时间,代表在此时生长速率最大;k为生长速率系数,曲线在过点(xc,a/e)时拥有最大斜率,即最大的生长速度,为
A×k/e 。1.2.3 发酵液代谢物前处理
精确移取100 µL发酵液至1.5 mL离心管中,加入400 µL提取液(甲醇:乙腈=1:1(v:v)),含0.02 mg/mL的内标(L-2-氯苯丙氨酸);涡旋混匀30 s后,低温超声提取30 min(5 ℃, 40 kHz),将样品静置于−20 ℃,30 min;离心15 min(13000 g,4 ℃),移取上清液,氮气吹干;加入100 μL复溶液(乙腈:水=1:1)复溶;涡旋混匀30 s,低温超声萃取5 min(5 ℃,40 kHz);离心10 min(13000 g,4 ℃),移取上清液至带内插管的进样小瓶中进行液相色谱-串联质谱分析。另外,每个样本分别移取20 µL上清液混合后作为质控样本。
1.2.4 液相色谱-串联质谱条件
色谱条件:色谱柱为ACQUITY UPLC HSS T3(100 mm×2.1 mm, 1.8 µm; Waters, Milford, USA);流动相A为95%水+5%乙腈(含0.1%甲酸),流动相B为47.5%乙腈+47.5%异丙醇+5%水(含0.1%甲酸),进样量为2 μL,柱温为40 ℃,流速0.4 mL/min,梯度洗脱程序:0~3.5 min,0%~24.5%B;3.5~5.5 min,24.5%~65%B;5.5~7.4 min,65%~100%B;7.4~7.6 min,100%~51.5%B;7.6~7.8 min,51.5%~0%B;7.8~10 min,0%B。
样品经电喷雾电离,分别采用正、负离子扫描模式采集质谱信号。质谱条件如下:扫描质荷比范围70~1050,鞘气流速50 L/min,辅助气流速13 L/min,加热温度425 ℃,毛细管温度325 ℃,喷雾电压±3500 V,碰撞能40 eV。
1.2.5 非靶向代谢组学表达量数据预处理
根据样本或分组内缺失值的比例,进行数据过滤,去除每一组内>20%的缺失值;并用极小值方法对原始数据中的缺失值进行模拟填充;数据归一化方法为总和;设置质控样本(Quality Control, QC)中相对标准偏差值≤30%。
1.3 数据处理
利用IBM SPSS Statistics 26对数据进行显著性分析等处理,使用Origin Pro 2019 b对数据进行拟合和作图,利用美吉生物生信云平台对非靶向代谢组学数据进行整合分析。
利用scipy (Python)进行京都基因与基因组百科全书(Kyoto Encyclopedia of Genes and Genomes, KEGG)代谢通路富集、相关性分析、聚类分析、Vip分析;利用ropls (R packages)进行PCA分析及OPLS-DA分析,分析数据均使用均值中心化方法进行数据转换,差异检验方法为Student's t-test,除特殊说明外文中置信度均为0.95。
2. 结果与分析
2.1 双歧杆菌BB12代谢茯苓多糖的生长曲线模型及评价
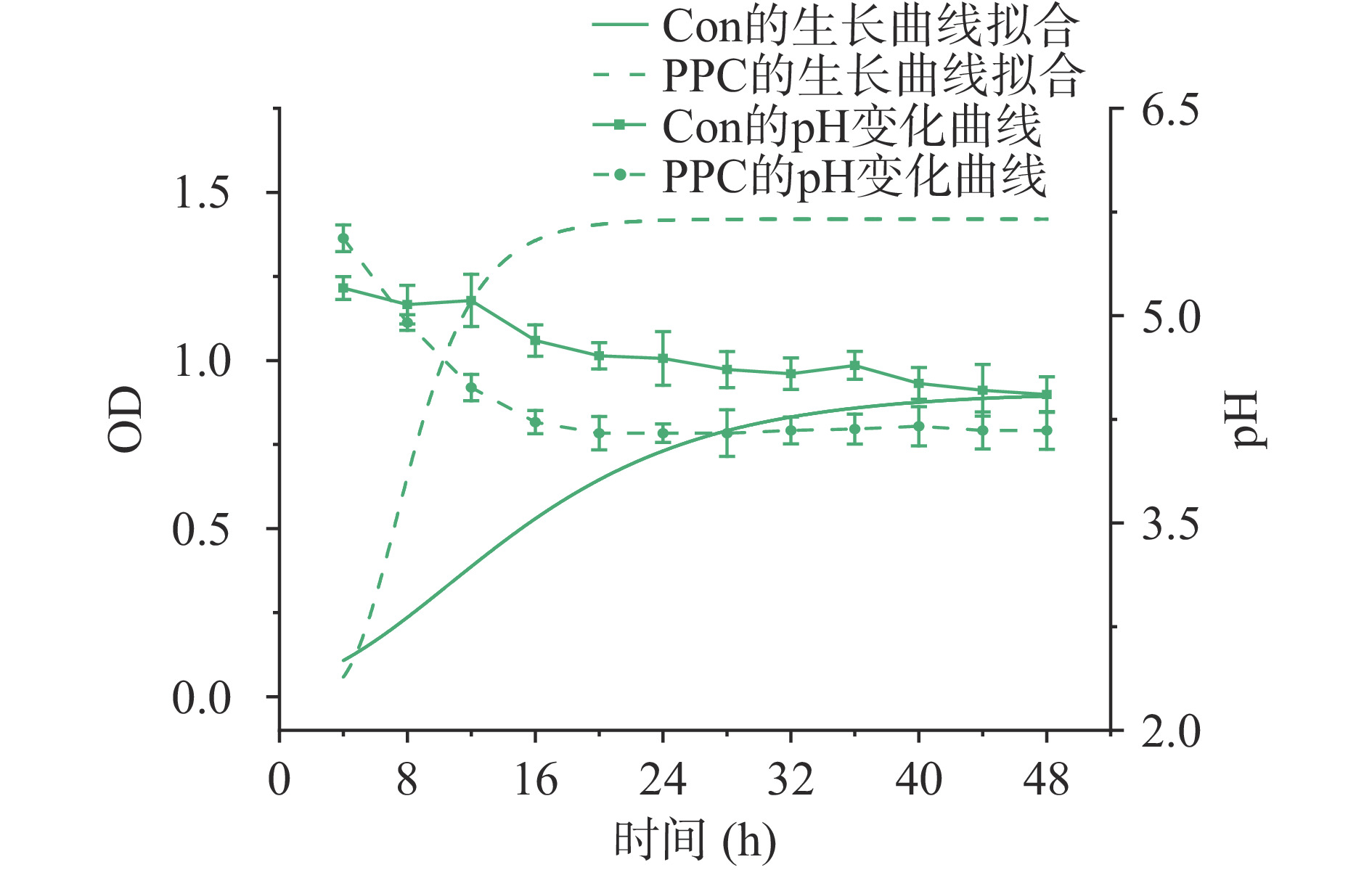
本实验以葡萄糖为唯一碳源的正常MRS肉汤培养基作为对照,用以比较双歧杆菌BB-12以茯苓多糖为唯一碳源时的生长曲线及发酵液的pH变化情况。双歧杆菌BB-12发酵葡萄糖的组合用Con表示(对照组,下同),双歧杆菌BB-12发酵茯苓多糖的组合用PPC表示(实验组,下同)。双歧杆菌BB-12随时间发酵葡萄糖和茯苓多糖的生长变化状况及pH变化如图1所示。表1给出了拟合模型的相关参数,模型拟合的决定系数R2均大于0.9,表明模型拟合效果较好。
![]() 图 1 双歧杆菌BB-12发酵葡萄糖和茯苓多糖的生长拟合曲线及pH变化曲线Figure 1. Growth fitting curve and pH change curve of glucose and polysaccharides of Poria cocos fermented by Bifidobacterium BB-12表 1 生长动力学方程非线性拟合相关参数Table 1. Parameters related to nonlinear fitting of growth dynamics equation
图 1 双歧杆菌BB-12发酵葡萄糖和茯苓多糖的生长拟合曲线及pH变化曲线Figure 1. Growth fitting curve and pH change curve of glucose and polysaccharides of Poria cocos fermented by Bifidobacterium BB-12表 1 生长动力学方程非线性拟合相关参数Table 1. Parameters related to nonlinear fitting of growth dynamics equation参数 Con PPC A 0.9065±0.0214 1.3571±0.0216 xc 10.5945±0.5079 8.9977±0.2519 k 0.1147±0.0108 0.5379±0.0809 R2 0.9888 0.9861 注:各参数意义见实验方法1.2.2。 结合图1和表1可知,双歧杆菌BB-12在以茯苓多糖为碳源的培养基中增殖迅速,细菌进入对数生长期的时间快于对照组(实验组xc值小于对照组xc),且进入稳定期后期望最大生长数量大于对照组(实验组A值大于对照组)。由此表明,实验组所添加的茯苓多糖相比对照组可有效增强双歧杆菌BB-12的代谢速率,并促进其增殖。
2.2 非靶向代谢组学样本相关性分析
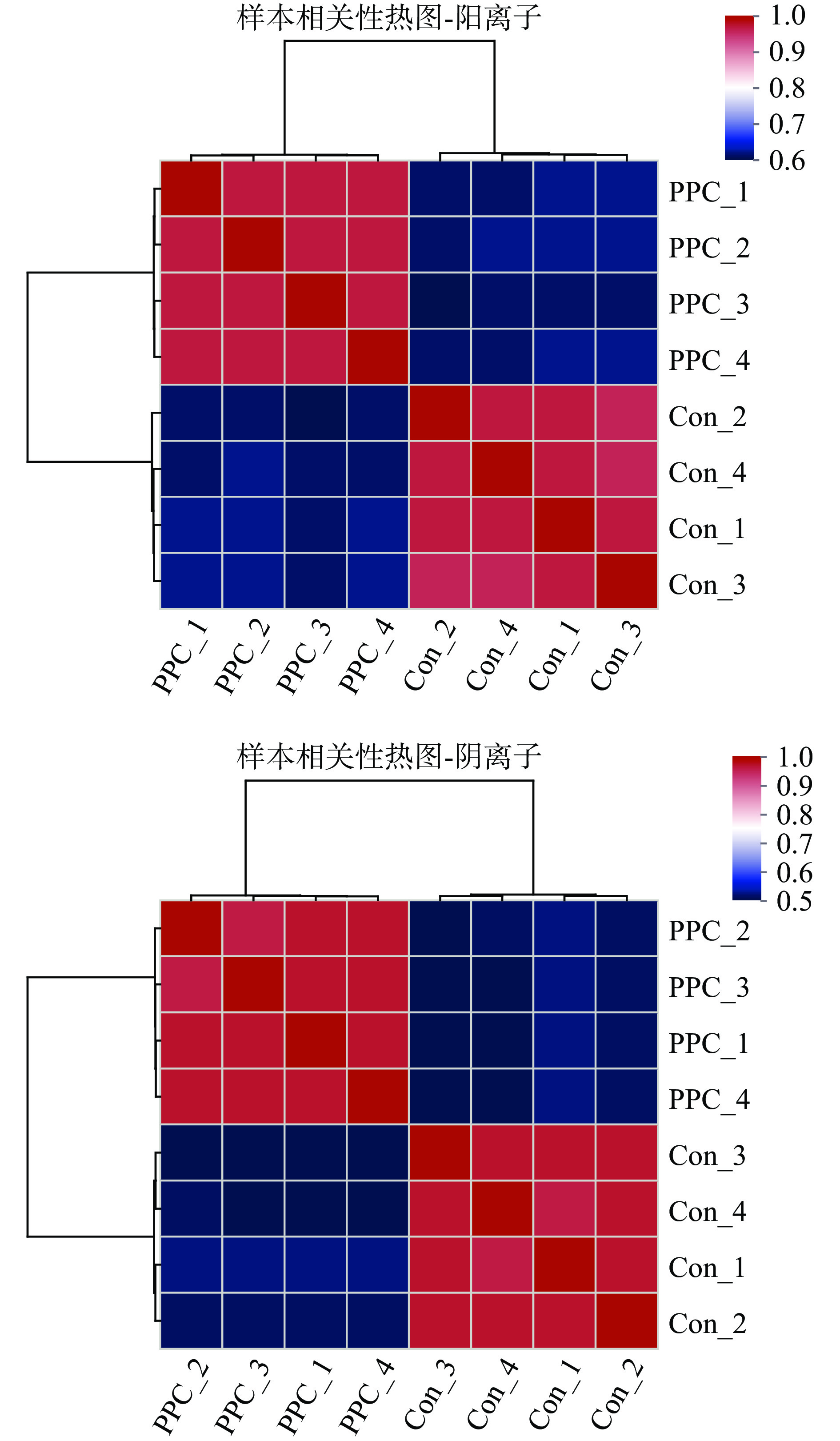
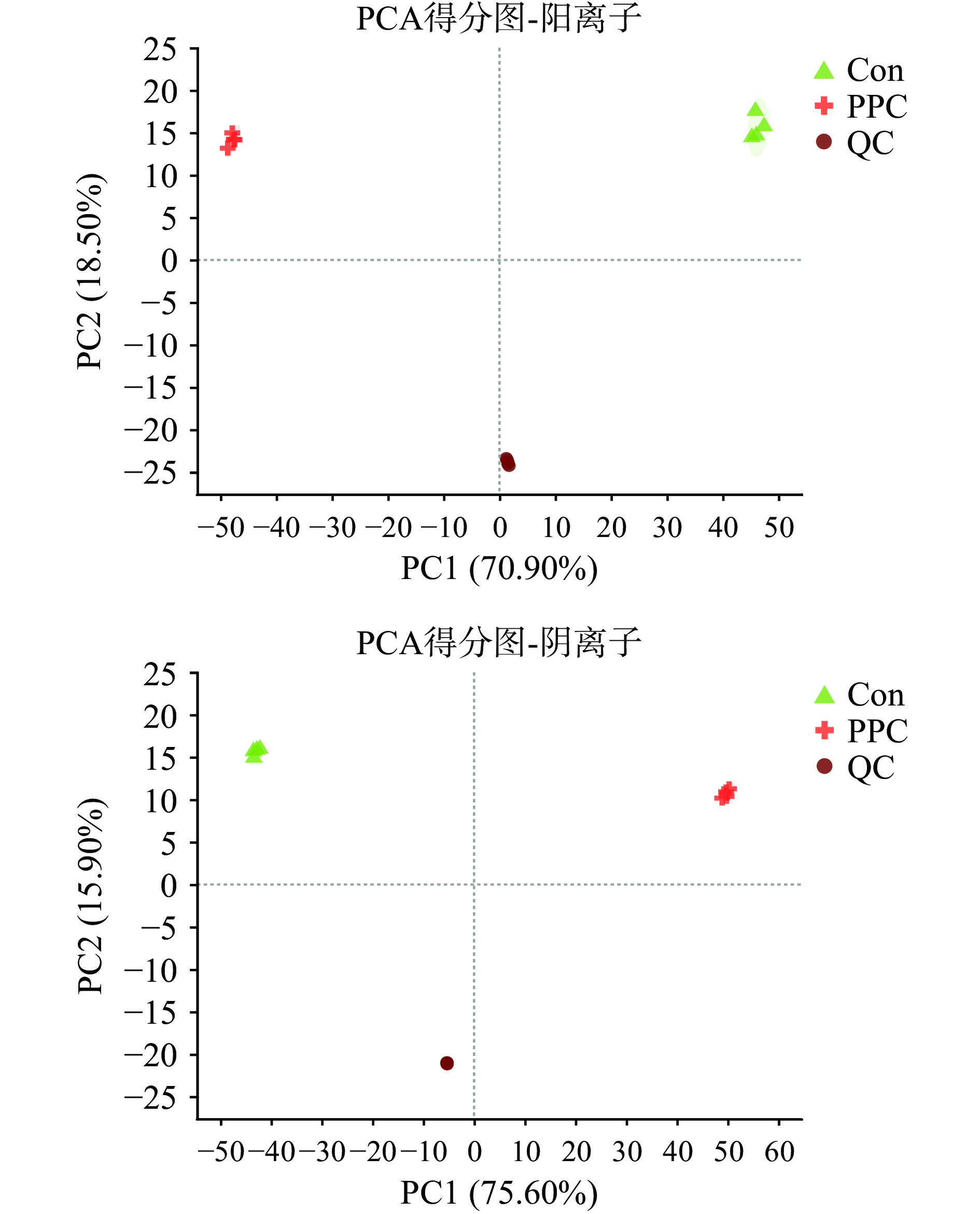
非靶向代谢组学样本相关性热图如图2所示。由图可知同组内样本相关性较高,即组内差异较小,反映样本之间存在的误差较小。图3分别为正、负离子模式下样本的PCA得分图。R2X(cum)表示累积的差异解释性,数值越接近1说明模型越好。图中正离子模式PC1的R2X(cum)为0.709,PC2的R2X(cum)为0.894,PC1与PC2对模型的贡献率为89.40%;负离子模式PC1的R2X(cum)为0.756,PC2的R2X(cum)为0.915,PC1与PC2对模型的贡献率为91.50%,主成分对模型贡献率较高,累积的差异解释性较好。
![]() 图 2 非靶向代谢组学样本相关性热图注:图中的右侧和下侧为样本名,图中每个格子表示两个样之间的相关性,不同颜色代表样本间相关系数的相对大小,聚类树枝的长度表示样本间相对距离的远近,同一枝上的样本中比较相似。Figure 2. Correlation heat map of non-targeted metabolomics samples
图 2 非靶向代谢组学样本相关性热图注:图中的右侧和下侧为样本名,图中每个格子表示两个样之间的相关性,不同颜色代表样本间相关系数的相对大小,聚类树枝的长度表示样本间相对距离的远近,同一枝上的样本中比较相似。Figure 2. Correlation heat map of non-targeted metabolomics samples根据图中结果可知,各组样本在图中位置分布相对集中,QC样本分布效果良好,即组内各样本差异不大,实验重复性好,随机误差较小,且模型对实际样本分布的解释性与准确性较好,可进一步对样本进行差异性分析。
2.3 差异代谢物的筛选与分析
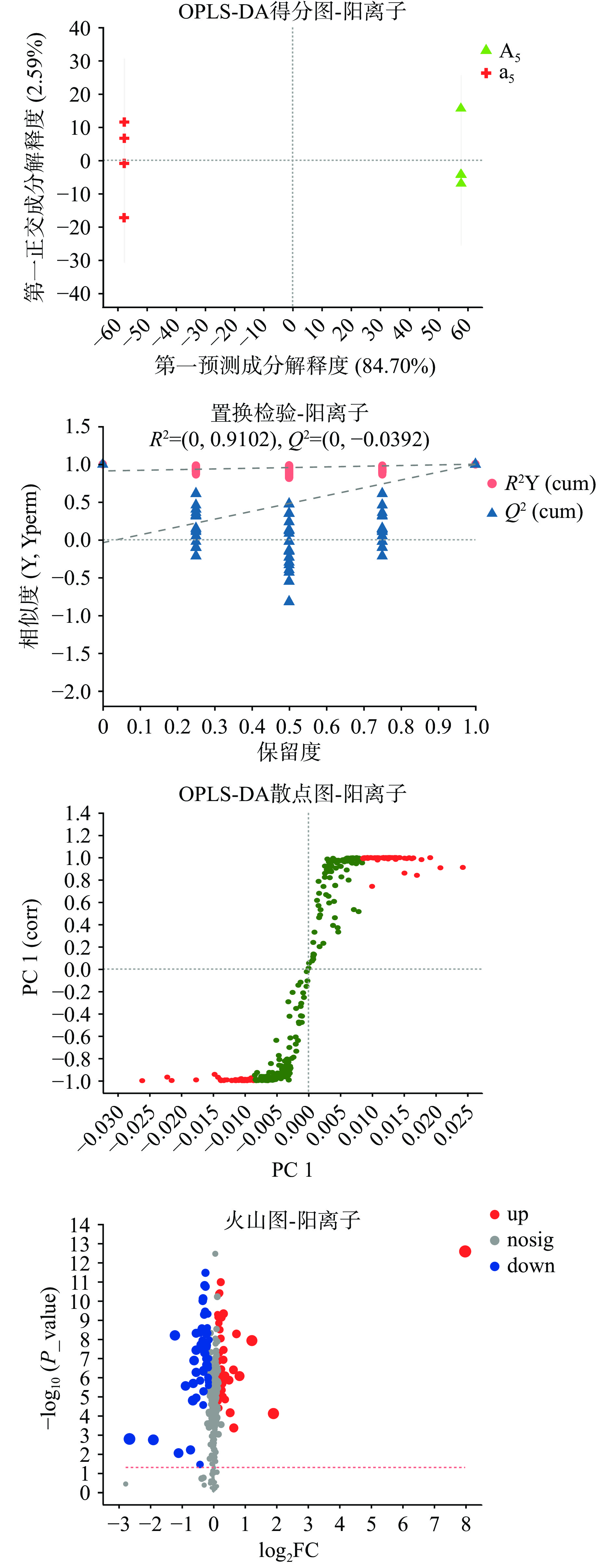
对于不同分组的组间差异比较,采用有监督的正交偏最小二乘回归模型的判别分析方法(Orthogonal Partial Least Squares Discriminant Analysis, OPLS-DA),并采用置换检验来标识监督性学习方法获得分类不是偶然的。通过这种方式,可以更好地区分组间差异,提高模型的有效性和解析能力。此外,采用OPLS-DA模型的VIP>1,P<0.05为筛选标准初步筛选出各组的差异代谢物进行下一步分析,并用S-plot与火山图对结果分布进行展示。
图4与图5分别为OPLS-DA模型建立结果的正离子模式和负离子模式展示图。结合图4、图5及表2数据,可见模型稳定可靠且预测能力较好。正、负离子模型的置换检验结果Q2(cum)截距值小于0.05,说明本实验建立的OPLS-DA模型未发生过拟合。结合数据库比对,共筛选出101个阳离子差异代谢物,80个阴离子差异代谢物。其中,有93个代谢物显著下调,88个代谢物显著上调。图6展示了VIP值前50的差异代谢物。
表 2 模型对X和Y矩阵的解释率及预测能力参数表Table 2. Parameters of the model's explanatory rate and predictive power for X and Y matrices参数 pos neg R2X(cum) 0.847 0.871 R2Y(cum) 1.000 1.000 Q2 0.999 0.999 注:R2X(cum)和R2Y(cum)表示所建模型对X和Y矩阵的累积解释率;Q2标示模型的预测能力,这3个指标越接近于1时表示模型越稳定可靠。 ![]() 图 6 双歧杆菌BB-12发酵葡萄糖和茯苓多糖的差异代谢物(Top50)注:左侧为代谢物聚类树状图,分支越近,说明样本内所有代谢物的表达模式越接近;每列表示一个样本,下方为样本名称;每行表示一个代谢物,颜色表示该代谢物在该组样本中相对表达量的大小,颜色梯度与数值大小的对应关系见梯度色块。右侧为代谢物VIP条形图,条形长度表示该代谢物对两组差异的贡献值,默认不小于1,值越大表示该代谢物在两组间差异越大。条形颜色表示代谢物在两组样本中差异显著性,即P_value值,P_value越小,-log10(P-value) 越大,颜色越深。右侧*代表P<0.05,**代表P<0.01,***代表P<0.001。Figure 6. Differential metabolites of fermented glucose and polysaccharides of Poria cocos by Bifidobacterium BB-12 (Top50)
图 6 双歧杆菌BB-12发酵葡萄糖和茯苓多糖的差异代谢物(Top50)注:左侧为代谢物聚类树状图,分支越近,说明样本内所有代谢物的表达模式越接近;每列表示一个样本,下方为样本名称;每行表示一个代谢物,颜色表示该代谢物在该组样本中相对表达量的大小,颜色梯度与数值大小的对应关系见梯度色块。右侧为代谢物VIP条形图,条形长度表示该代谢物对两组差异的贡献值,默认不小于1,值越大表示该代谢物在两组间差异越大。条形颜色表示代谢物在两组样本中差异显著性,即P_value值,P_value越小,-log10(P-value) 越大,颜色越深。右侧*代表P<0.05,**代表P<0.01,***代表P<0.001。Figure 6. Differential metabolites of fermented glucose and polysaccharides of Poria cocos by Bifidobacterium BB-12 (Top50)上述结果显示,双歧杆菌BB-12代谢茯苓多糖产生的代谢产物中,上调最显著的是一种异喹啉生物碱(Oxoglaucine),报道[16]称该物质具有较强的药理活性,有抗菌、抗炎、抗癌、修复免疫系统等诸多功能[17],CHEN等[18]实验证明它是一类可有效治疗乳腺癌的功能成分。其次上调的还有丹芝酸、芒柄花素、黄藤素、羟基积雪草酸(Madecassic acid)以及三萜类化合物等成分;羟基积雪草酸是积雪草中有效活性物质,已有的报道证明其具有抗氧化、抗抑郁、抗菌、调节免疫等生物活性功能[19],ZHANG等[20]将CT26结肠癌细胞注射到小鼠体内并用不同剂量的羟基积雪草酸进行治疗,发现羟基积雪草酸在体内可通过促使肿瘤细胞凋亡并增强免疫来抑制肿瘤生长;WON等[21]通过研究其对小鼠单核巨噬细胞白血病细胞RAW264.7的作用发现,羟基积雪草酸可通过调节NF-κB信号通路进而影响肿瘤坏死因子TNF-α、白细胞介素IL-1β、IL-6等物质的分泌,从而在体内达到抗炎的效果。黄藤素(Palmatine)同样具有抗炎、抗菌、提高免疫能力等作用[22],并且对多种癌细胞系具有抗癌活性[23],它还可通过诱导胰岛素的分泌来达到降低血糖的效果[24],并降低血液中总胆固醇和甘油三酯含量,提高高密度脂蛋白的含量[25]。TAKAYAMA等[26]及PARK等[25]分别通过实验证实了双歧杆菌可以调节血脂代谢、治疗糖尿病等潜在功效,该代谢物的产生可能是双歧杆菌发挥上述功效的原因之一。芒柄花素(Formononetin)是一种异黄酮类化合物,近年来因其抗氧化、抗癌、抗炎等生物学特性而广受关注[27]。许多研究表明刺芒柄花素具有抑制肿瘤细胞增殖的潜力,如通过多种信号通路诱导肿瘤细胞凋亡等[28-29],此外,在医学领域,这种活性化合物在预防和治疗多种疾病方面显示出潜力,包括肥胖和神经神经退化性疾病性疾病等[18]。三萜类化合物((3beta,17alpha,23R)-17,23-Epoxy-3,29-dihydroxy-27-norlanost-8-ene-15,24-dione)是由数个异戊二烯去掉羟基后首尾相连构成的具有多种生物活性的物质,如灵芝三萜有抗氧化、抗菌、抗肿瘤以及免疫调节活性等[30];此外在差异代谢物中差异含量较大的还有丹芝酸E(Ganolucidic acid E)、丹芝酸B(Ganolucidic acid B)和丹芝酸C(Ganolucidic acid C),丹芝酸类也是一种具有较强生理功效的三萜类化合物,有研究证明丹芝酸具有保护肝脏活性[31]和治疗HIV-1的潜在功能[32];上述分析结果显示,这些代谢产物可能是茯苓在人体内发挥功效的关键因素之一,而肠道内的菌群尤其是双歧杆菌对其代谢可能是该物质生成的关键前提。
显著下调的代谢物有同型胱氨酸、酒石酸、千里光宁碱、胆酸和葡萄糖基胺等。其中,同型胱氨酸和同型半胱氨酸的升高被认为是冠状动脉、脑动脉和周围血管发生动脉粥样硬化的独立危险因素[33]。张丽杰等[34]也通过临床实验证实口服双歧杆菌可以降低胆固醇、甘油三酯以及低密度脂蛋白胆固醇的含量,对治疗动脉粥样硬化和心脑血管疾病有一定意义。该代谢物的显著下调说明茯苓多糖可以更加充分的发挥双歧杆菌对这些疾病的预防作用。酒石酸是一种具有肌肉毒性的物质,它可以抑制细胞中苹果酸的生产,而苹果酸是有氧代谢的重要中间产物;千里光宁碱(Senecionine),是一种有毒的生物碱,可通过调节细胞内钙离子的分布从而表达肝脏毒性[35]。这一结果表明,下调这些代谢产物可能与双歧杆菌BB-12发酵茯苓多糖比发酵葡萄糖表现出更好的生长性能及减少体内炎症等有关。
2.4 KEGG差异代谢通路的富集
KEGG代谢通路图表示物种代谢的反应、相互作用网络以及遗传、环境信息处理和其他细胞过程等。这些通路不仅提供生化物质相互转化所有可能的代谢途径,还包含对催化各步反应的酶的全面注解等[36]。
通过与KEGG数据库比对分析,发现差异代谢物共富集到61条代谢通路,其中,显著富集17条(图7)。富集最显著的前三个通路为ABC转运蛋白通路、亚油酸代谢、半乳糖代谢。ABC转运蛋白(ATP—binding cassette transpoter)是一类庞大而古老的跨膜运输蛋白家族,在生物体内参与多种物质的转运积累、有害物质解毒、气孔调节、植物防御等生理活动。亚油酸是一种人体必需的不饱和脂肪酸,能降低血液胆固醇,预防动脉粥样硬化。许多文献如[37-38]都表明一些双歧杆菌可以产生亚油酸,RHEE等[39]的研究也证实双歧杆菌在预防动脉硬化方面有一定作用。与对照组相比,实验组中富集到该通路的代谢物均有显著上调,这表明茯苓多糖可能促进双歧杆菌产生亚油酸及相关衍生物,这条通路可能也是双歧杆菌改善体内血脂代谢的潜在机制之一。对半乳糖的代谢是双歧杆菌较其他微生物所表现出较强的代谢功能之一,该代谢通路的显著富集也印证了这一点。
![]() 图 7 KEGG差异代谢通路气泡图注:横坐标为富集率;纵坐标为KEGG通路。图中气泡的大小代表该通路中富集到代谢集中代谢物数量的多少,气泡的颜色表示不同富集显著性P值的大小。Figure 7. KEGG differential metabolic pathway bubble diagram
图 7 KEGG差异代谢通路气泡图注:横坐标为富集率;纵坐标为KEGG通路。图中气泡的大小代表该通路中富集到代谢集中代谢物数量的多少,气泡的颜色表示不同富集显著性P值的大小。Figure 7. KEGG differential metabolic pathway bubble diagram富集因子最高的通路为胆碱能药物和抗胆碱能药物途径(Cholinergic and anticholinergic drugs,Pathway ID:map07220),富集因子达到0.5。该通路中鉴定到了一个关键代谢物安普托品(Amprotropine),在该代谢通路中安普托品的前体物质是阿托品,阿托品在胆碱能神经节后纤维效应器处,可与递质的乙酰胆碱争夺受体,主流研究认为乙酰胆碱含量增多与阿尔兹海默病的症状改善显著相关。在该条通路中也显示,这一物质是抗帕金森药物的主要原料之一。现有的研究结果也报道了双歧杆菌对帕金森病[40]以及阿尔兹海默症[41]的治疗有促进作用。因此,这条通路可能是双歧杆菌在肠道内发挥脑-肠轴功效的关键通路之一。此外,在该通路中安普托品是治疗肠稳态失衡的前体物质,该代谢物在实验组中较对照组显著上调,表明茯苓多糖较葡萄糖可以更好地发挥双歧杆菌在维持肠稳态中的重要益生功能。
本研究结合OPLS-DA差异代谢物模型及KEGG差异代谢通路,共筛选出一种三萜类化合物((3beta,17alpha,23R)-17,23-Epoxy-3,29-dihydroxy-27-norlanost-8-ene-15,24-dione)、芒柄花素、安普托品、同型胱氨酸、酒石酸、丹芝酸E、丹芝酸B、丹芝酸C、异喹啉生物碱、黄藤素、千里光宁碱、羟基积雪草酸共12种具有相关生理活性的关键差异代谢物,并以箱体图形式(图8)展现对照组与实验组之间含量丰度差异。
通过对差异代谢物的相关分析发现,双歧杆菌BB-12代谢茯苓多糖产生了许多三萜类、黄酮类物质,且这些物质大多具有较强的生物活性,如抗炎、抗癌、调节机体免疫等。而这些活性恰好是茯苓多糖在动物实验和临床研究中显示的结果。由此提示,上述代谢产物可能是茯苓发挥与疾病调控相关功能的原因之一。此外,通过对差异代谢物的富集得到了几条重要的代谢通路,这些代谢通路也可能是双歧杆菌与茯苓多糖在体内发挥相关益生功效的关键途径。
3. 结论
本文通过非靶向代谢组学技术,从小分子代谢物层面比较分析了动物双歧杆菌乳双歧亚种BB-12发酵茯苓多糖和葡萄糖的代谢产物之间的差异,这些代谢物以及所对应的代谢通路不仅与茯苓多糖和双歧杆菌的抗菌、抗炎、调节机体免疫、改善肠道菌群等益生功能密切相关,且与肠道菌群与机体各部位之间如肠-脑轴、肠-肝轴等的平衡与调节有关。由此,从科学的角度进一步证实了我国传统中药材茯苓多糖发挥益生功效的部分机制,也为今后探究双歧杆菌与益生元之间相关性研究建立一种科学有效的模型,并为未来疾病的靶向预防与调控,以及个性化饮食营养干预奠定了一定的理论基础。
-
![]()
图 1 双歧杆菌BB-12发酵葡萄糖和茯苓多糖的生长拟合曲线及pH变化曲线
Figure 1. Growth fitting curve and pH change curve of glucose and polysaccharides of Poria cocos fermented by Bifidobacterium BB-12
![]()
图 2 非靶向代谢组学样本相关性热图
注:图中的右侧和下侧为样本名,图中每个格子表示两个样之间的相关性,不同颜色代表样本间相关系数的相对大小,聚类树枝的长度表示样本间相对距离的远近,同一枝上的样本中比较相似。
Figure 2. Correlation heat map of non-targeted metabolomics samples
![]()
图 6 双歧杆菌BB-12发酵葡萄糖和茯苓多糖的差异代谢物(Top50)
注:左侧为代谢物聚类树状图,分支越近,说明样本内所有代谢物的表达模式越接近;每列表示一个样本,下方为样本名称;每行表示一个代谢物,颜色表示该代谢物在该组样本中相对表达量的大小,颜色梯度与数值大小的对应关系见梯度色块。右侧为代谢物VIP条形图,条形长度表示该代谢物对两组差异的贡献值,默认不小于1,值越大表示该代谢物在两组间差异越大。条形颜色表示代谢物在两组样本中差异显著性,即P_value值,P_value越小,-log10(P-value) 越大,颜色越深。右侧*代表P<0.05,**代表P<0.01,***代表P<0.001。
Figure 6. Differential metabolites of fermented glucose and polysaccharides of Poria cocos by Bifidobacterium BB-12 (Top50)
![]()
图 7 KEGG差异代谢通路气泡图
注:横坐标为富集率;纵坐标为KEGG通路。图中气泡的大小代表该通路中富集到代谢集中代谢物数量的多少,气泡的颜色表示不同富集显著性P值的大小。
Figure 7. KEGG differential metabolic pathway bubble diagram
表 1 生长动力学方程非线性拟合相关参数
Table 1 Parameters related to nonlinear fitting of growth dynamics equation
参数 Con PPC A 0.9065±0.0214 1.3571±0.0216 xc 10.5945±0.5079 8.9977±0.2519 k 0.1147±0.0108 0.5379±0.0809 R2 0.9888 0.9861 注:各参数意义见实验方法1.2.2。  下载: 导出CSV
下载: 导出CSV
表 2 模型对X和Y矩阵的解释率及预测能力参数表
Table 2 Parameters of the model's explanatory rate and predictive power for X and Y matrices
参数 pos neg R2X(cum) 0.847 0.871 R2Y(cum) 1.000 1.000 Q2 0.999 0.999 注:R2X(cum)和R2Y(cum)表示所建模型对X和Y矩阵的累积解释率;Q2标示模型的预测能力,这3个指标越接近于1时表示模型越稳定可靠。
下载: 导出CSV
-
[1] 王悦, 田双双, 刘晓谦, 等. 茯苓多糖的提取、结构及药理作用研究进展[J]. 世界中医药,2021,16(17):2548−2555. [WANG Y, TIAN S S, LIU X Q, et al. Research progress on extraction, structures and pharmacological activities of Poria cocos polysaccharides[J]. World Chinese Medicine,2021,16(17):2548−2555. doi: 10.3969/j.issn.1673-7202.2021.17.006 [2] RÍOS J L. Chemical constituents and pharmacological properties of Poria cocos[J]. Planta Medica,2011,77(7):681−691. doi: 10.1055/s-0030-1270823
[3] KHAN I, HUANG G, LI X, et al. Mushroom polysaccharides from Ganoderma lucidum and Poria cocos reveal prebiotic functions[J]. Journal of Functional Foods,2018,41:191−201. doi: 10.1016/j.jff.2017.12.046
[4] GAO S, LAI C K M, CHEUNG P C K. Nondigestible carbohydrates isolated from medicinal mushroom sclerotia as novel prebiotics[J]. International Journal of Medicinal Mushrooms,2009,11(1):1−8.
[5] JIANG Y, FAN L. The effect of Poria cocos ethanol extract on the intestinal barrier function and intestinal microbiota in mice with breast cancer[J]. Journal of Ethnopharmacology,2021,266:113456. doi: 10.1016/j.jep.2020.113456
[6] SONG K Y, JIANG Z Y, YAN Q C, et al. Experimental study on the effect of Radix codonopsis and Poria on intestinal flora in mice[J]. The Chinese Journal of Clinical Pharmacology,2011,27(2):142−145.
[7] JUNGERSEN M, WIND A, JOHANSEN E, et al. The science behind the probiotic strain Bifidobacterium animalis subsp. lactis BB-12®[J]. Microorganisms,2014,2(2):92−110. doi: 10.3390/microorganisms2020092
[8] MÄTTÖ J, FONDÉN R, TOLVANEN T, et al. Intestinal survival and persistence of probiotic Lactobacillus and Bifidobacterium strains administered in triple-strain yoghurt[J]. International Dairy Journal,2006,16(10):1174−1180. doi: 10.1016/j.idairyj.2005.10.007
[9] SAVARD P, LAMARCHE B, PARADIS M E, et al. Impact of Bifidobacterium animalis subsp. lactis BB-12 and, Lactobacillus acidophilus LA-5-containing yoghurt, on fecal bacterial counts of healthy adults[J]. International Journal of Food Microbiology,2011,149(1):50−57. doi: 10.1016/j.ijfoodmicro.2010.12.026
[10] CHOURAQUI J P, VAN EGROO L D, FICHOT M C. Acidified milk formula supplemented with Bifidobacterium lactis: Impact on infant diarrhea in residential care settings[J]. Journal of Pediatric Gastroenterology and Nutrition,2004,38(3):288−292. doi: 10.1097/00005176-200403000-00011
[11] WANG K Y, LI S N, LIU C S, et al. Effects of ingesting Lactobacillus and Bifidobacterium containing yogurt in subjects with colonized Helicobacter pylori[J]. The American Journal of Clinical Nutrition,2004,80(3):737−741.
[12] COMMANE D M, SHORTT C T, SILVI S, et al. Effects of fermentation products of pro-and prebiotics on trans-epithelial electrical resistance in an in vitro model of the colon[J]. Nutrition and Cancer,2005,51(1):102−109. doi: 10.1207/s15327914nc5101_14
[13] MARTINS F S, SILVA A A, VIEIRA A T, et al. Comparative study of Bifidobacterium animalis, Escherichia coli, Lactobacillus casei and Saccharomyces boulardii probiotic properties[J]. Archives of Microbiology,2009,191(8):623−630. doi: 10.1007/s00203-009-0491-x
[14] ZWIETERING M H, JONGENBURGER I, ROMBOUTS F M, et al. Modeling of bacterial growth curve[J]. Applied and Environmental Microbiology,1990,56(6):1875−1881. doi: 10.1128/aem.56.6.1875-1881.1990
[15] KATHLEEN M C TJØRVE, E TJØRVE. The use of Gompertz models in growth analyses, and new Gompertz-model approach: An addition to the Unified-Richards family[J]. Plos One,2017,12(6):e0178691. doi: 10.1371/journal.pone.0178691
[16] CHEN Z F, SHI Y F, LIU Y C, et al. TCM active ingredient oxoglaucine metal complexes: Crystal structure, cytotoxicity, and interaction with DNA[J]. Inorganic Chemistry,2012,51(4):1998−2009. doi: 10.1021/ic200443p
[17] REMICHKOVA M, DIMITROVA P, PHILIPOV S, et al. Toll-like receptor-mediated anti-inflammatory action of glaucine and oxoglaucine[J]. Fitoterapia,2009,80(7):411−414. doi: 10.1016/j.fitote.2009.05.016
[18] CHEN T, CHEN H, ZHANG L, et al. Analysis of oxoglaucine in the treatment of breast cancer based on network pharmacology and bioinformatics[C]//E3S Web of Conferences. EDP Sciences, 2021, 271: 03078.
[19] YANG B, XU Y, HU Y, et al. Madecassic acid protects against hypoxia-induced oxidative stress in retinal microvascular endothelial cells via ROS-mediated endoplasmic reticulum stress[J]. Biomedicine & Pharmacotherapy,2016,84:845−852.
[20] ZHANG H, ZHANG M, TAO Y, et al. Madecassic acid inhibits the mouse colon cancer growth by inducing apoptosis and immunomodulation[J]. J BUON,2014,19:372−376.
[21] WON J H, SHIN J S, PARK H J, et al. Anti-inflammatory effects of madecassic acid via the suppression of NF-κB pathway in LPS-induced RAW 264.7 macrophage cells[J]. Planta Medica,2010,76(3):251−257. doi: 10.1055/s-0029-1186142
[22] ZHANG L, LI J, MA F, et al. Synthesis and cytotoxicity evaluation of 13-n-Alkyl berberine and palmatine analogues as anticancer agents[J]. Molecules,2012,17(10):11294−11302. doi: 10.3390/molecules171011294
[23] PATEL M B, MISHRA S. Hypoglycemic activity of alkaloidal fraction of Tinospora cordifolia[J]. Phytomedicine,2011,18(12):1045−1052. doi: 10.1016/j.phymed.2011.05.006
[24] MA H, HU Y, ZOU Z, et al. Antihyperglycemia and antihyperlipidemia effect of protoberberine alkaloids from Rhizoma coptidis in HepG2 cell and diabetic KK-Ay mice[J]. Drug Dev Res,2016,77(4):163−170. doi: 10.1002/ddr.21302
[25] PARK Y H, KIM J G, SHIN Y W, et al. Effects of Lactobacillus acidophilus 43121 and a mixture of Lactobacillus casei and Bifidobacterium longum on the serum cholesterol level and fecal sterol excretion in hypercholesterolemia-induced pigs[J]. Bioscience, Biotechnology, and Biochemistry,2008,72(2):595−600. doi: 10.1271/bbb.70581
[26] TAKAYAMA F, TAKI K, NIWA T. Bifidobacterium in gastro-resistant seamless capsule reduces serum levels of indoxyl sulfate in patients on hemodialysis[J]. American Journal of Kidney Diseases,2003,41(3):S142−S145. doi: 10.1053/ajkd.2003.50104
[27] CHULWON K, SEOK-GEUN L, YANG W M, et al. Formononetin-induced oxidative stress abrogates the activation of STAT3/5 signaling axis and suppresses the tumor growth in multiple myeloma preclinical model[J]. Cancer Letters,2018,431:123−141. doi: 10.1016/j.canlet.2018.05.038
[28] PARK S, BAZER F W, LIM W, et al. The O-methylated isoflavone, formononetin, inhibits human ovarian cancer cell proliferation by sub G0/G1 cell phase arrest through PI3K/AKT and ERK1/2 inactivation[J]. Journal of Cellular Biochemistry, 2018.
[29] DUTRA J M, ESPITIA P, BATISTA R A . Formononetin: Biological effects and uses-A review[J]. Food Chemistry, 2021, 359(5):129975.
[30] 刘维, 虎虓真, 朱莉, 等. 灵芝三萜的研究与应用进展[J]. 食品科学,2019,40(5):309−315. [LIU W, HU X Z, ZHU L, et al. Recent progress in research and application of Ganoderma lucidum triterpenoids[J]. Food Science,2019,40(5):309−315. doi: 10.7506/spkx1002-6630-20180201-021 [31] LIU L Y, WANG H Q, LIU C, et al. Triterpenoids of Ganoderma sessile and their hepatoprotective activities[J]. Natural Product Research and Development,2017,29(4):584.
[32] KANG D, MUTAKIN M, LEVITA J. Computational study of triterpenoids of Ganoderma lucidum with aspartic protease enzymes for discovering HIV-1 and plasmepsin inhibitors[J]. International Journal of Chemistry,2015,7(1):62.
[33] TAYLOR J R, LLOYD M, et al. The association of elevated plasma homocyst (e) ine with progression of symptomatic peripheral arterial disease[J]. Journal of Vascular Surgery,1991,13(1):128−136.
[34] 张丽杰, 高峰, 王春艳, 等. 双歧杆菌活菌降低老年人血脂的临床研究[J]. 中国卫生产业,2012,9(23):67. [ZHANG L J, GAO F, WANG C Y, et al. A clinical study on the effect of live bifidobacteria on reducing blood lipids in the elderly[J]. China Health Industry,2012,9(23):67. [35] GRIFFIN D S, SEGALL H J. Effects of the pyrrolizidine alkaloid senecionine and the alkenals trans-4-OH-hexenal and trans-2-hexenal on intracellular calcium compartmentation in isolated hepatocytes[J]. Biochemical Pharmacology,1989,38(3):391−397. doi: 10.1016/0006-2952(89)90377-8
[36] KANEHISA M. KEGG for representation and analysis of molecular networks involving diseases and drugs[J]. Nucleic Acids Research,2010,38(Database issue):D355−D360.
[37] COAKLEY M, ROSS R P, NORDGREN M, et al. Conjugated linoleic acid biosynthesis by human-derived Bifidobacterium species[J]. Journal of Applied Microbiology,2003,94(1):138−145. doi: 10.1046/j.1365-2672.2003.01814.x
[38] GORISSEN L, RAES K, WECKX S, et al. Production of conjugated linoleic acid and conjugated linolenic acid isomers by Bifidobacterium species[J]. Applied Microbiology and Biotechnology,2010,87(6):2257−2266. doi: 10.1007/s00253-010-2713-1
[39] RHEE Y K, HAN M J, CHOI E C, et al. Hypocholesterolemic activity of Bifidobacteria isolated from a healthy Korean[J]. Archives of Pharmacal Research,2002,25(5):681−684. doi: 10.1007/BF02976944
[40] ISHII T, FURUOKA H, KAYA M, et al. Oral administration of probiotic Bifidobacterium breve improves facilitation of Hippocampal memory extinction via restoration of aberrant higher induction of neuropsin in an MPTP-induced mouse model of Parkinson’s disease[J]. Biomedicines,2021,9(2):167. doi: 10.3390/biomedicines9020167
[41] KOBAYASHI Y, SUGAHARA H, SHIMADA K, et al. Therapeutic potential of Bifidobacteriumbreve strain A1 for preventing cognitive impairment in Alzheimer’s disease[J]. Scientific Reports,2017,7(1):1−10. doi: 10.1038/s41598-016-0028-x


 下载:
下载:












计量
- 文章访问数: 233
- HTML全文浏览量: 208
- PDF下载量: 45
