


Isolation of Enterobacter hohensis 4-2-1 from Human Intestine and Its Fermentation Effect on Lycium Barbarum Polysaccharide
-
摘要: 为了阐明肠道细菌在枸杞多糖吸收过程中的作用,本论文主要针对1株从人体肠道粪便中分离到的细菌进行研究,并将该菌株用于枸杞多糖发酵,并以发酵中细菌总数、pH、多糖含量、还原糖含量和分子量为指标反映发酵特性,初步阐明了枸杞多糖的消化和吸收。结果表明,通过形态学观察和分子生物学鉴定,该菌株为肠道单菌霍氏肠杆菌4-2-1(Enterobacter Hoechnii),为革兰氏阴性菌,且该菌在发酵到12 h细菌总数达到最大,发酵后,培养基的pH由7.98变为7.75、多糖含量由682 μg/mL变为205.7 μg/mL、还原糖含量由225 μg/mL变为91.32 μg/mL,分子量由8.02 kDa变为3.44 kDa。结果发现,枸杞多糖在经过霍氏肠杆菌发酵后,多糖含量下降为原来的30.2%左右,还原糖含量下降为40.6%左右,分子量下降为42.9%,说明肠道菌能发酵利用枸杞多糖,研究为进一步研究多糖和肠道菌的相互作用提供依据。Abstract: In order to investigate the role of intestinal bacteria in the absorption of Lycium barbarum polysaccharides (LBP), a strain of bacteria was isolated from human intestinal feces, and the strain was applied in the fermentation of LBP. The total number of bacteria, pH, polysaccharide content, reducing sugar content and molecular weight in the fermentation were measured and analysed to initially elucidate the digestion and absorption of LBP. The results showed that the strain was Enterobacter hormaechei through morphological and molecular biology identification, marked 4-2-1, which was a Gram-negative bacterium, and that it reached its maximum total bacterial count at 12 h. After fermentation, the pH of the medium changed from 7.98 to 7.75, the polysaccharide content changed from 682 μg/mL to 205.7 μg/mL, the reducing sugar content changed from 225 μg/mL to 91.32 μg/mL, and the molecular weight changed from 8.02 to 3.44 kDa, the content of LBP decreased to about 30.2%, the content of reducing sugar decreased to about 40.6% and the molecular weight decreased to 42.9%. It indicated that the intestinal bacteria could ferment and utilize LBP, which would provide a basis for further study on the interaction between LBP and intestinal bacteria.
-
Keywords:
- Lycium barbarum polysaccharide /
- Enterobacter hormaechei /
- screening /
- fermentation
-
宁夏枸杞(Lycium barbarum L.)别名茄果,枸杞子,是我国备受喜爱的药食同源食物[1],其中含有多种活性成分,主要包括多糖、多酚、类胡萝卜素、萜烯类、有机酸以及多种微量元素[2]。而多糖又是枸杞中最主要的活性成分之一,在免疫调节、抗氧化、抗疲劳、护肝等方面贡献颇大[3-7]。枸杞多糖与疾病的相互作用已成为关注的热点。有研究表明,枸杞多糖不仅可以维持肠道微生物群的平衡,改善非酒精性肝病[8],而且能够调节肠道菌群,减少压力因素对后代情绪伤害的影响[9]。
人体肠道是一个大型的发酵、消化场所[10]。其中人体肠道菌群组成复杂、数量巨大,包括拟杆菌属、肠杆菌属、芽孢杆菌属等。霍氏肠杆菌属于肠杆菌科,是存在与人和动物肠道内的正常菌株之一,现在国内对霍氏肠杆菌的研究较少,汪芳芳[11]发现优化霍氏肠杆菌A20培养条件之后,叶黄素的降解率提高了20%左右,增加了叶黄素的商业价值,同时霍氏肠杆菌WM1可以发酵碳水化合物产生青霉素酶,并在检测牛奶中青霉素的残留发挥一定作用[12]。而人肠道菌群也不是固定不变的,与很多因素都有密切关系,比如年龄、环境、性别、饮食等多种因素,其中饮食被认为是改变肠道菌群最直接、最有效的方法之一[13]。有研究表明,碳水化合物是人类饮食的主要成分,但其中的植物多糖不能被口腔、胃液吸收,只有被分解成小分子物质才能达到其益生元作用,而肠道微生物能够定植在远端肠道分解复杂碳水化合物[14-15],Fu等[16]人对鼠尾藻多糖的体外发酵研究中发现,鼠尾藻多糖只有被结肠微生物发酵后才能对人体发挥作用,亚麻籽多糖在被人类粪便微生物水解利用后才能有助于成为促进结肠的益生元[17]。枸杞多糖是一种水溶性多糖,为含有多种微量元素和氨基酸的蛋白多糖,具有重要生理活性[18]。而目前对枸杞多糖的发酵作用大多研究主要集中在粪便菌群的发酵上[19],对单菌和枸杞多糖之间的作用了解甚少。因此明确对枸杞多糖发挥作用的肠道单菌具有重要意义。
本研究从为了阐明肠道细菌在人体吸收枸杞多糖过程中的作用,通过形态学观察,生理生化实验以及16S rRNA基因对其进行鉴定,筛选出一株肠道单菌株,最后将该单菌应用到枸杞多糖发酵中,通过对枸杞多糖发酵指标的研究,以揭示枸杞多糖在人体内的作用机制,为之后相关方面的研究提供一定的借鉴和参考。
1. 材料与方法
1.1 材料与仪器
人体肠道粪便 联系3位健康志愿者,要求志愿者无消化系统疾病,3个月内未服用抗生素或益生菌;枸杞,购自宁夏银川市中宁;枸杞多糖 由本实验室制备得到。细菌基因组DNA提取试剂盒 北京索莱宝科技有限公司;苯酚、浓硫酸 国药集团化学试剂有限公司,琼脂糖 北京擎科生物科技有限公司。
BXM-30R压力蒸汽灭菌器 上海博讯实业有限公司;超净工作台 北京东联哈尔仪器制造有限公司;V-5100紫外分光光度计 上海精科实业有限公司;LRH-250生化培养箱 广东省医疗器械厂;凝胶成像系统 Eppendorf公司;梯度PCR仪 Eppendorf公司;LB培养基 青岛高科技工业园海生物技术有限公司;琼脂粉 北京索莱宝科技有限公司;SpectraMax iD3多功能酶标仪 美谷分子仪器上海有限公司;S210 pH计 上海仪电科学仪器股份有限公司。
1.2 实验方法
1.2.1 培养基的配制
无碳源SM液体培养基:氯化钾4.5 g/L、氯化钠4.5 g/L、碳酸氢钠1.5 g/L、磷酸二氢钾0.5 g/L、磷酸氢二钾0.5 g/L、硫酸镁0.4 g/L、氯化铵0.3 g/L、碱性微量元素(Na2SeO3 0.087 g/L,Na2WO4 0.165 g/L,Na2MoO4 0.121 g/L,NaOH 2 g/L)1 mL/L、酸性微量元素(FeCl2、ZnCl2、CuCl2、HCl)1 mL/L,高温高压115 ℃灭菌15 min后,按照500:1的比例加入CaCl2母液,按照1000:1的比例加入维生素溶液(维生素B3、维生素B6、维生素B2、维生素B1),配制固体培养基所需琼脂粉为20 g/L。
1.2.2 人粪便样品预处理
称取一定质量新鲜人类粪便,在超净工作台中加入无菌PBS进行涡旋,500 r/min离心5 min,吸上清至灭完菌的离心管中,再加入无菌PBS旋涡,500 r/min离心5 min,再次吸上清至灭完菌的Ep管中,得到粪便悬液。粪便悬液一部分用于实验,剩余部分加入无菌厌氧甘油,分装,保存在−80 ℃,备用[19]。
1.2.3 人体粪便中菌的富集
取一定量粪便悬液,500 r/min离心5 min,弃去上清,用PBS洗涤两次后,离心重悬,按照合适的体积比,接种至LB液体培养基中,37 ℃摇床培养48 h。
1.2.4 肠道菌分离筛选
将1.2.3富集的样品用PBS溶液进行稀释至10−3~10−7梯度,在LB固体培养基中进行涂布,放置37 ℃恒温培养箱中进行培养24~36 h。挑选培养基中不同颜色、形状和大小的菌株至LB液体培养基,37 ℃摇床培养18~24 h。将液体培养基中得到的菌株在LB固体培养基中反复划线,已保证得到纯度较高的单菌,将上述得到的单菌落以50%甘油进行保菌,并保存于−80 ℃冰箱中。
1.2.5 菌株发酵能力的比较
将经过反复划线得到的两株单菌按照1:50的比例加入含有枸杞多糖的SM液体培养基中,放置在37 ℃摇床中摇床培养72 h,测量发酵液中多糖含量的变化。
1.2.6 菌株鉴定
将1.2.4中得到对枸杞多糖发酵能力较好的菌株4-2-1进行鉴定。根据《常见细菌系统鉴定手册》[20]和《伯杰氏细菌系统分类学手册》[21]对分离出的细菌进行细菌形态和生理生化鉴定。
采用16S rRNA基因进行分子生物学鉴定,以4-2-1菌株的基因组为模板进行PCR扩增,正反向引物为8F:5'-AGAGTTTGATCCTGGCTCAG-3'/1492R:5'-GGTTACCTTGTTACGACTT-3'。PCR反应体系(50 μL)为:模板DNA 1 μL,KOD酶1 μL,10×KOD buffer 5 μL,dNTP(2 mmol/L)5 μL,引物(F+R)3 μL,MgSO4(25 mmol/L) 3 μL,ddH2O 22 μL。PCR扩增参数设置为:94 ℃预变性2 min,98 ℃变性10 s,55 ℃退火30 s,68 ℃延将。得到的PCR扩增产物进行1%(质量分数)琼脂糖凝胶电泳。确认聚合酶反应扩增片段,若PCR扩增产物在1500 bp左右,则可送至上海生工进行测序,将该16S rDNA序列递交NCBI进行BLAST同源序列检索,比较筛出菌株与已知菌株的系统发育关系和系统地位,如果同源性大于99%,则确认为同一类细菌,借助Mega X软件建立系统发育树。
1.2.7 霍氏肠杆菌4-2-1对枸杞多糖发酵的影响
将枸杞多糖(2 mg/mL)加入无碳源的SM固体培养基中,使得枸杞多糖成为培养基中的唯一碳源,放置37 ℃恒温培养箱中对菌株4-2-1培养至有明显菌落。并接种至含枸杞多糖的无碳源SM液体培养基中,放置37 ℃摇床中培养18 h,培养后吸取1 mL的菌液,的以3000 r/min进行离心3 min,收集菌体并用PBS稀释至1 mL,按照1:50的比例将霍氏肠杆菌接种至以枸杞多糖为唯一碳源的SM液体培养基中进行摇床培养,动态监测发酵过程中细菌生长含量(OD600)、pH的变化、多糖含量、还原糖含量以及分子量变化。
1.2.7.1 发酵过程中菌株生长曲线测定
将菌株按照1:50的比例加入含有枸杞多糖为唯一碳源的SM液体培养基中,放置在37 ℃摇床中摇床培养,分别在0、6、12、24、48 h取样,用紫外分光光度计测每个时间点的OD600,实验在相同条件下重复三次。
1.2.7.2 发酵过程中pH测定
将菌株加入含有枸杞多糖为唯一碳源的的SM液体培养基中,放置在37 ℃摇床中摇床培养,分别在0、6、12、24、48 h取样,用pH计测每个时间点的pH,以按照1:50的比例接种霍氏肠杆菌4-2-1的SM液体培养基作为对照组,实验在相同条件下重复三次。
1.2.7.3 发酵过程中多糖含量测定
硫酸-苯酚法测定发酵液中多糖含量,参考Masuko等[22]方法略作修改。取各个时间段的发酵液,13000 r/min离心3 min,取上清,分别吸取0.3 mL不同时间点发酵样品溶液和葡萄糖标准溶液加入0.2 mL的6%苯酚溶液,震荡摇匀后迅速加入1 mL浓硫酸,摇匀加塞后置于沸水浴放置20 min,取出后用凉水冷却10 min;用酶标仪进行测定,在波长为490 nm处测定吸光度,以葡萄糖浓度X为横坐标(µg/mL),OD490nm为纵坐标,绘制标准曲线,并根据标准曲线Y=0.02200X+0.04749,R2=0.9997计算样品中总碳水化合物含量。
1.2.7.4 发酵中还原糖含量测定
采用二硝基水杨酸(DNS)法测定发酵液中还原糖,参照牟佳红等[23]方法略作修改。分别吸取0.4 mL不同时间点发酵样品溶液和葡萄糖标准溶液,加入DNS试剂0.8 mL,摇匀后置于恒温混匀仪100 ℃加热5 min,取出后置于冰水混合物中冷却至室温,用酶标仪进行测定,在波长为540 nm处测定吸光度,以葡萄糖浓度X为横坐标(mg/mL),D540nm为纵坐标,绘制标准曲线,并根据标准曲线Y=1.54300X+0.04671,R2=0.9991计算样品还原糖含量含量。
1.2.7.5 发酵过程中多糖分子量测定
分子量的测定参照唐雨薇[24]方法略作修改。发酵产物使用80%的乙醇进行4 ℃过夜沉淀,并用0.22 μm滤膜过滤。采用Wasters-2424蒸发光检测器测定多糖的分子量,检测条件:色谱柱:GE水溶性凝胶柱(7.8 mm×300 mm);柱温:30 ℃;流速:0.6 mL/min;流动相:纯化水;雾化器模式:加热(90%功率,36 ℃);载气:N2;气流压力:0.28 MPa;柱温:30 ℃;进样量:10 μL。
1.3 数据处理
实验均重复三次取平均值,实验结果均用平均值±标准差来表示,采用Mega X软件建立系统发育树,用统计学分析软件SPSS进行显著性分析,Excel 2019进行数据统计并分析,由Graph Pad Prism 8软件进行实验数据作图。
2. 结果与分析
2.1 发酵能力较优菌株的筛选
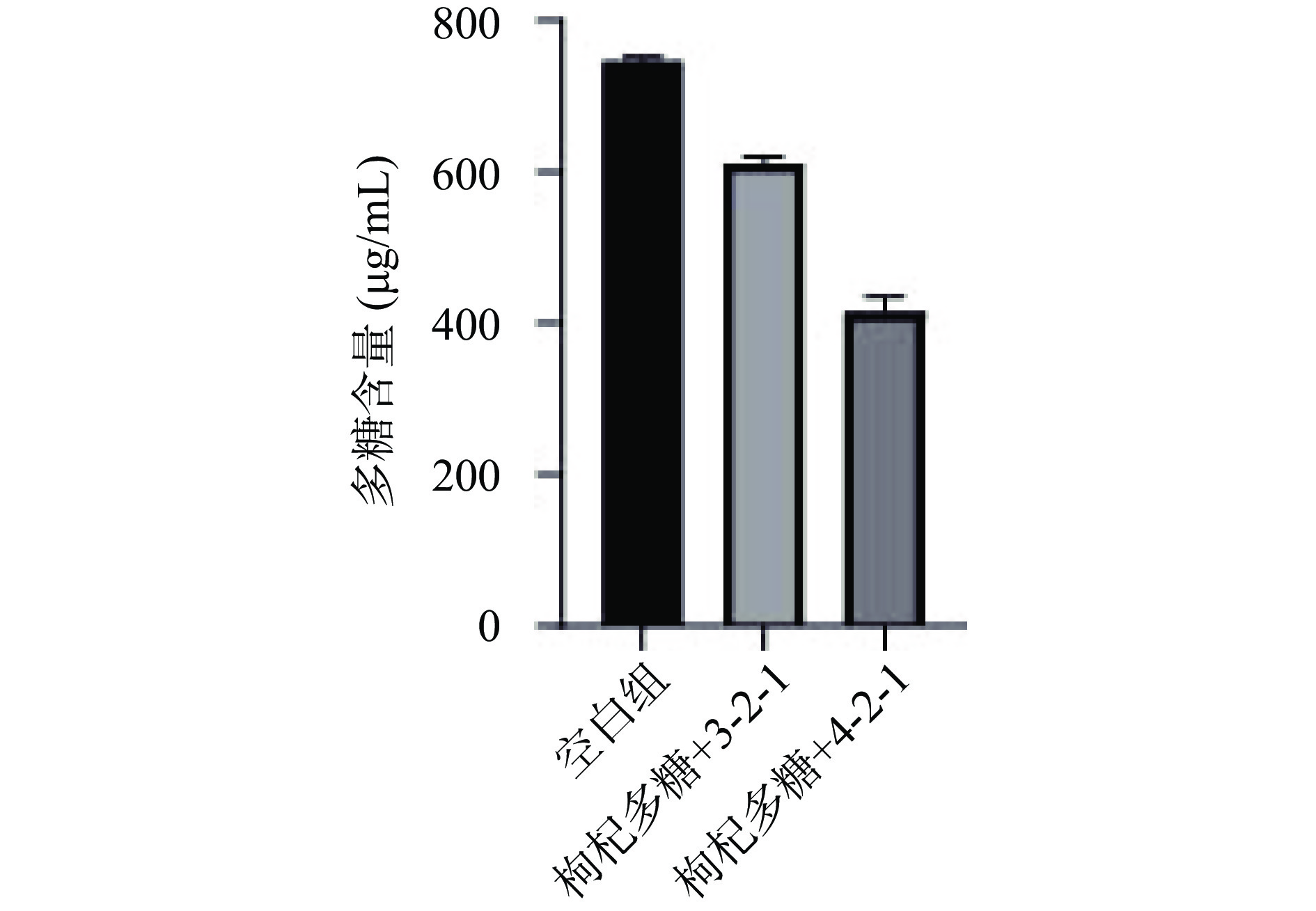
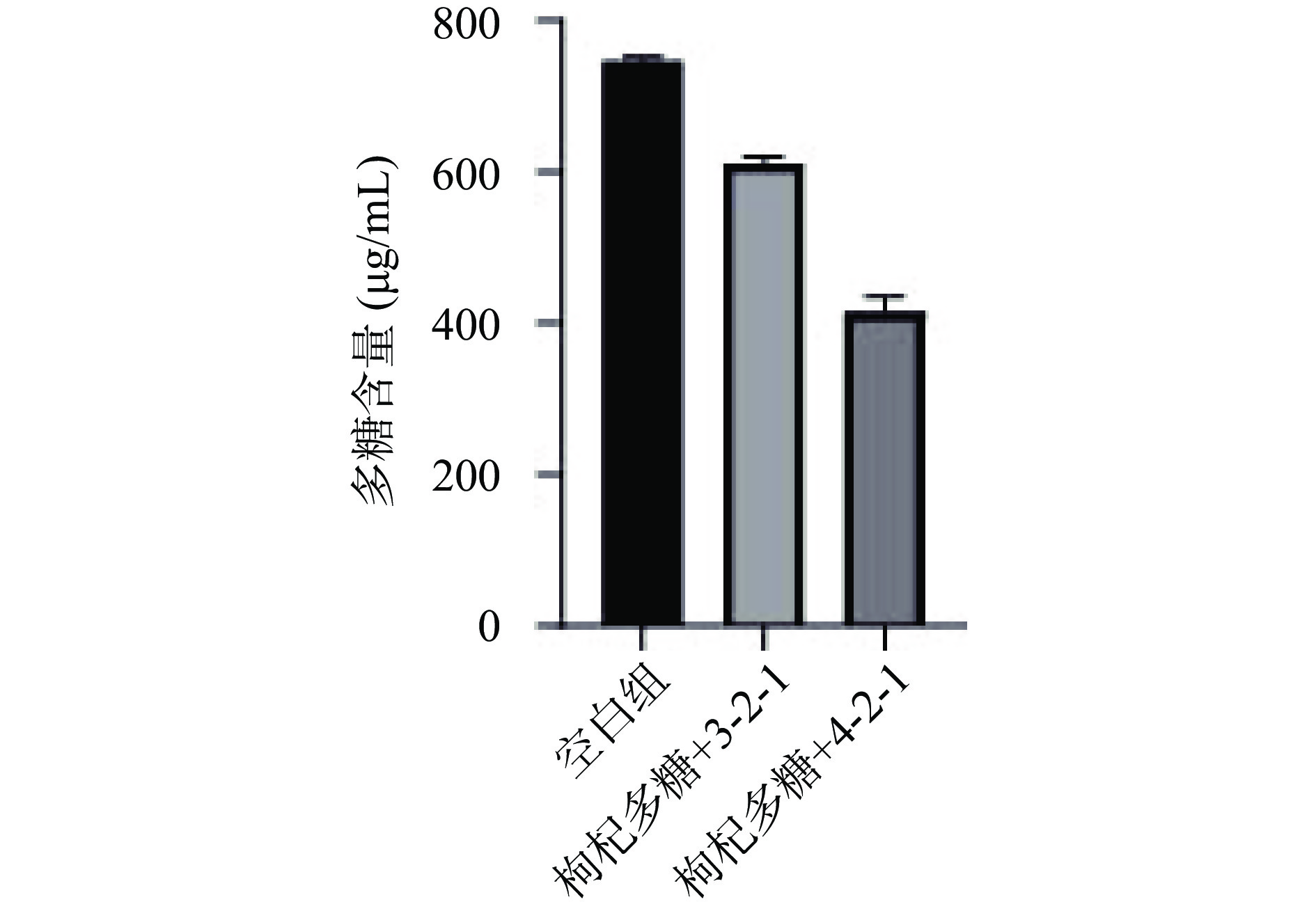
通过反复划线可得纯度较高的两株细菌,将菌株3-2-1和菌株4-2-1接种至含枸杞多糖的无碳源SM液体培养基中进行培养,测定培养基中多糖含量的变化,无碳源SM培养基作为对照组。由图1可知,发酵72 h后,与对照组相比,两个发酵组多糖含量明显降低,说明这两株菌均可以利用枸杞多糖,并且菌株4-2-1的发酵能力强于3-2-1,因此选择发酵能力较优菌株4-2-1进行后续实验。
![]() 图 1 菌株筛选过程中多糖含量的变化Figure 1. Changes in polysaccharide content during strain screening
图 1 菌株筛选过程中多糖含量的变化Figure 1. Changes in polysaccharide content during strain screening2.2 人肠道菌株的鉴定
2.2.1 形态学鉴定
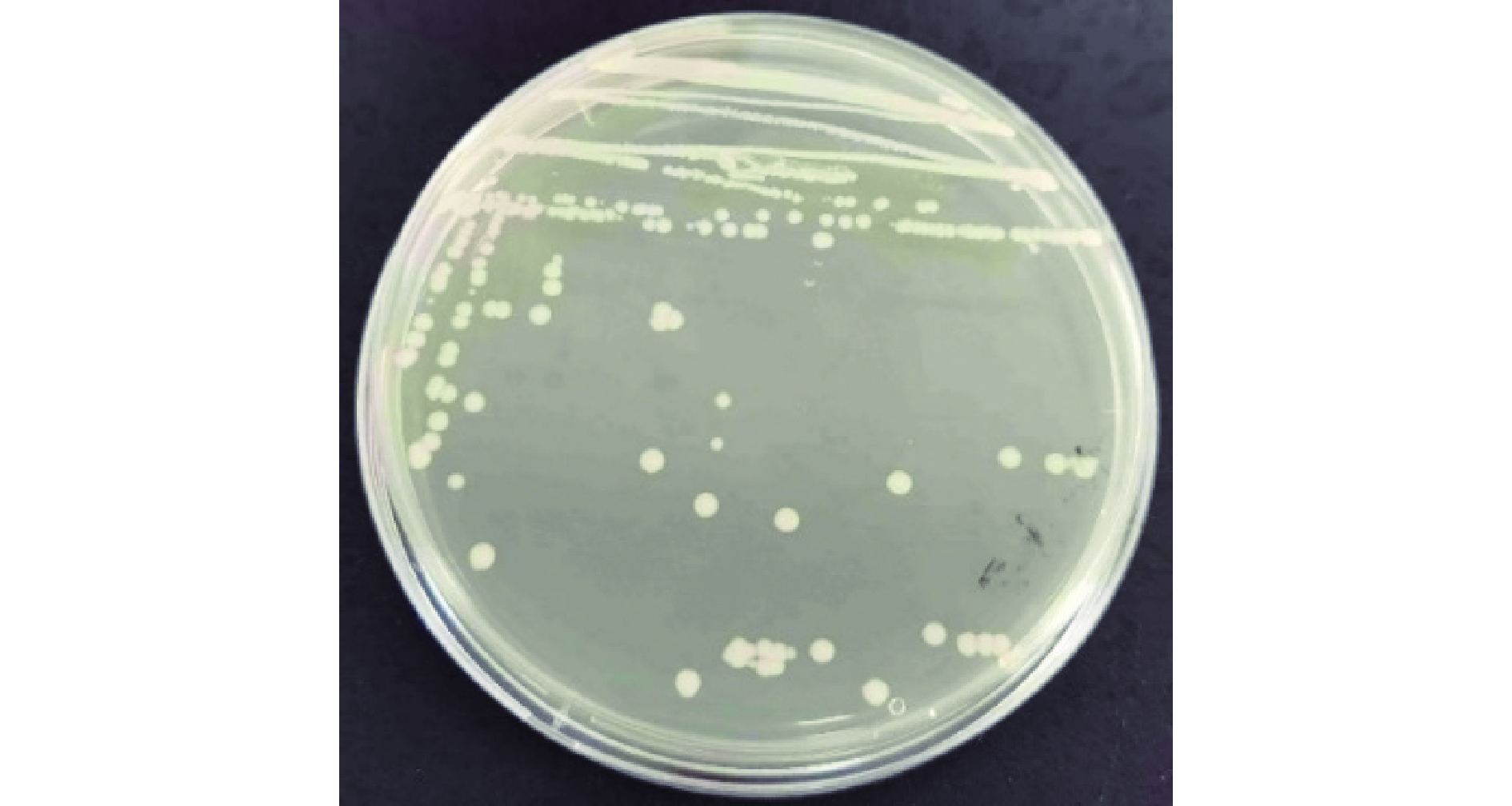
采用三区划线法将4-2-1在LB固体培养基上划线。通过对4-2-1菌株形态学观察,发现菌株4-2-1在LB培养基平板上生长情况如图2,圆形,边缘透明,中间呈乳白色,有光泽,略隆起,边缘整齐;对4-2-1菌株进行革兰氏染色后发现,该菌周身鞭毛运动,无芽孢,无夹膜,通过革兰氏染色初步判定4-2-1为革兰氏阴性菌,在显微镜下成杆状。
2.2.2 16S rRNA扩增结果
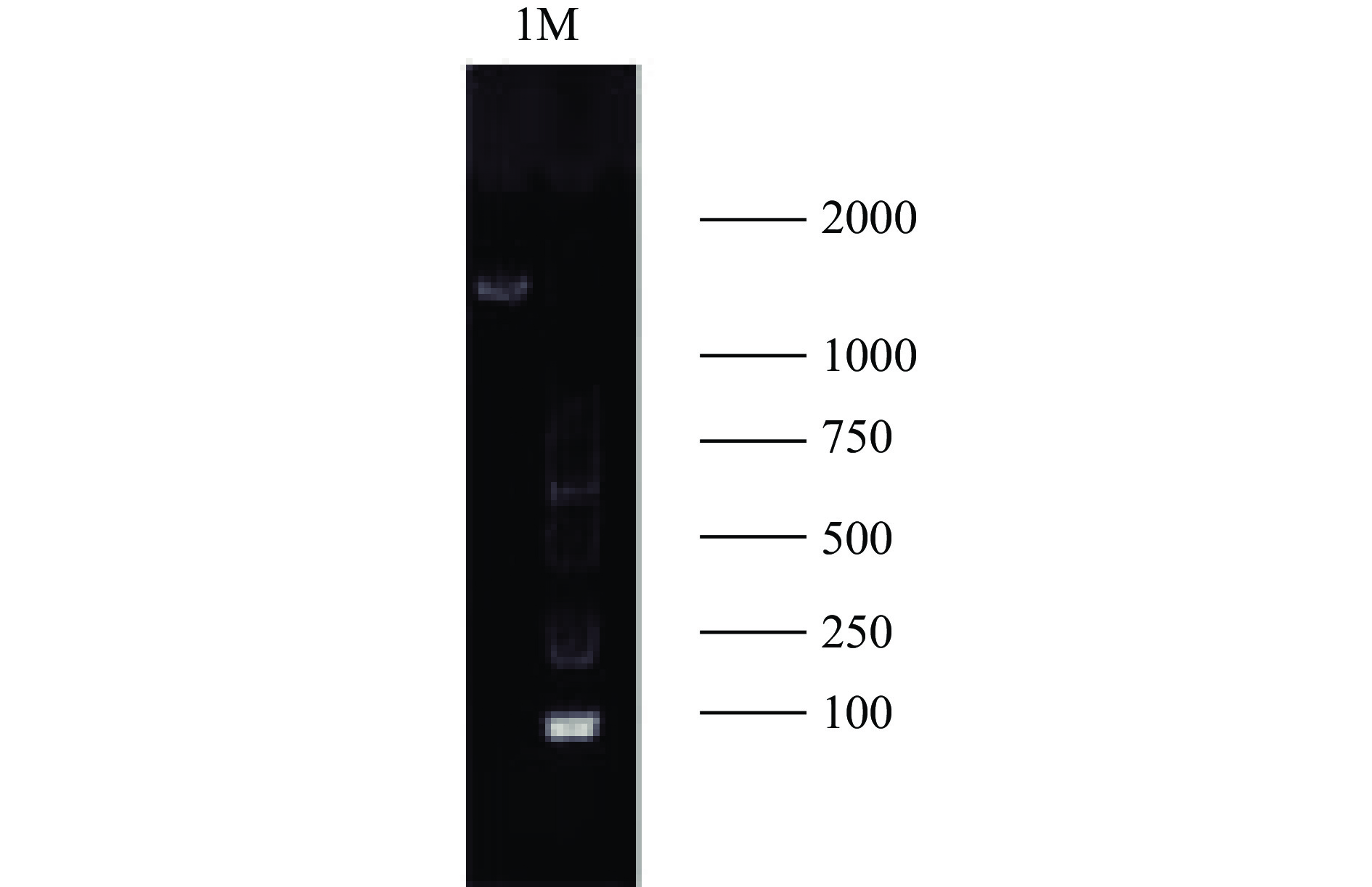
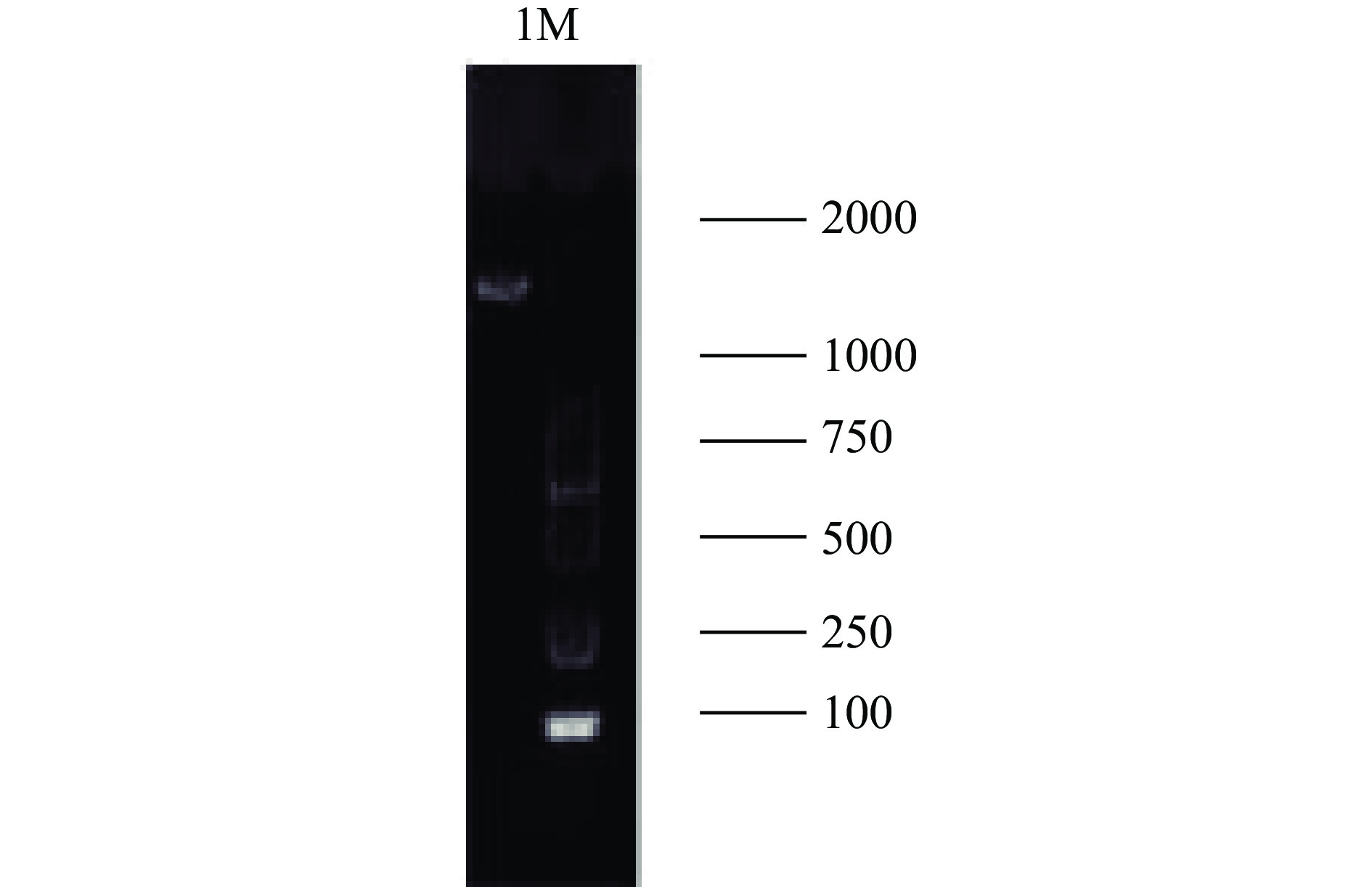
利用引物对所提取的基因组DNA进行扩增,所扩增的结果如图3所示,在1500 bp左右出现了目的条带,条带整齐,表明PCR产物可用于测序分析。
![]() 图 3 1%琼脂凝胶电泳图注:M:2000 DNAMaker;1:所提菌株基因组。Figure 3. Agarose gel electrophoresis diadram
图 3 1%琼脂凝胶电泳图注:M:2000 DNAMaker;1:所提菌株基因组。Figure 3. Agarose gel electrophoresis diadram2.2.3 4-2-1系统进化树分析
用Snap Gene Viewer软件对4-2-1测序序列进行拼接,借助NCBI中的BLAST和数据库中已知菌株的16S rRNA 基因进行序列比对,用相似度最高的菌株来确定4-2-1的种属状况。结果表明,菌4-2-1的16S rRNA 基因序列与数据库中已知霍氏肠杆菌的16S rRNA基因的相似性大于99%。为了进一步明确该菌株与已知菌株的亲缘关系及分类地位,构建系统发育树,结果显示该菌株与Enterobacter hormaechei KK736258.1在系统发育树聚为一支,系统发育树如图4所示。
![]() 图 4 菌株4-2-1的16S rRNA基因序列系统发育树Figure 4. Phylogenetic tree of Enterobacter 16S rRNA gene sequence of strain 4-2-1
图 4 菌株4-2-1的16S rRNA基因序列系统发育树Figure 4. Phylogenetic tree of Enterobacter 16S rRNA gene sequence of strain 4-2-12.3 对枸杞多糖发酵的影响
2.3.1 发酵筛选及菌株生长特性
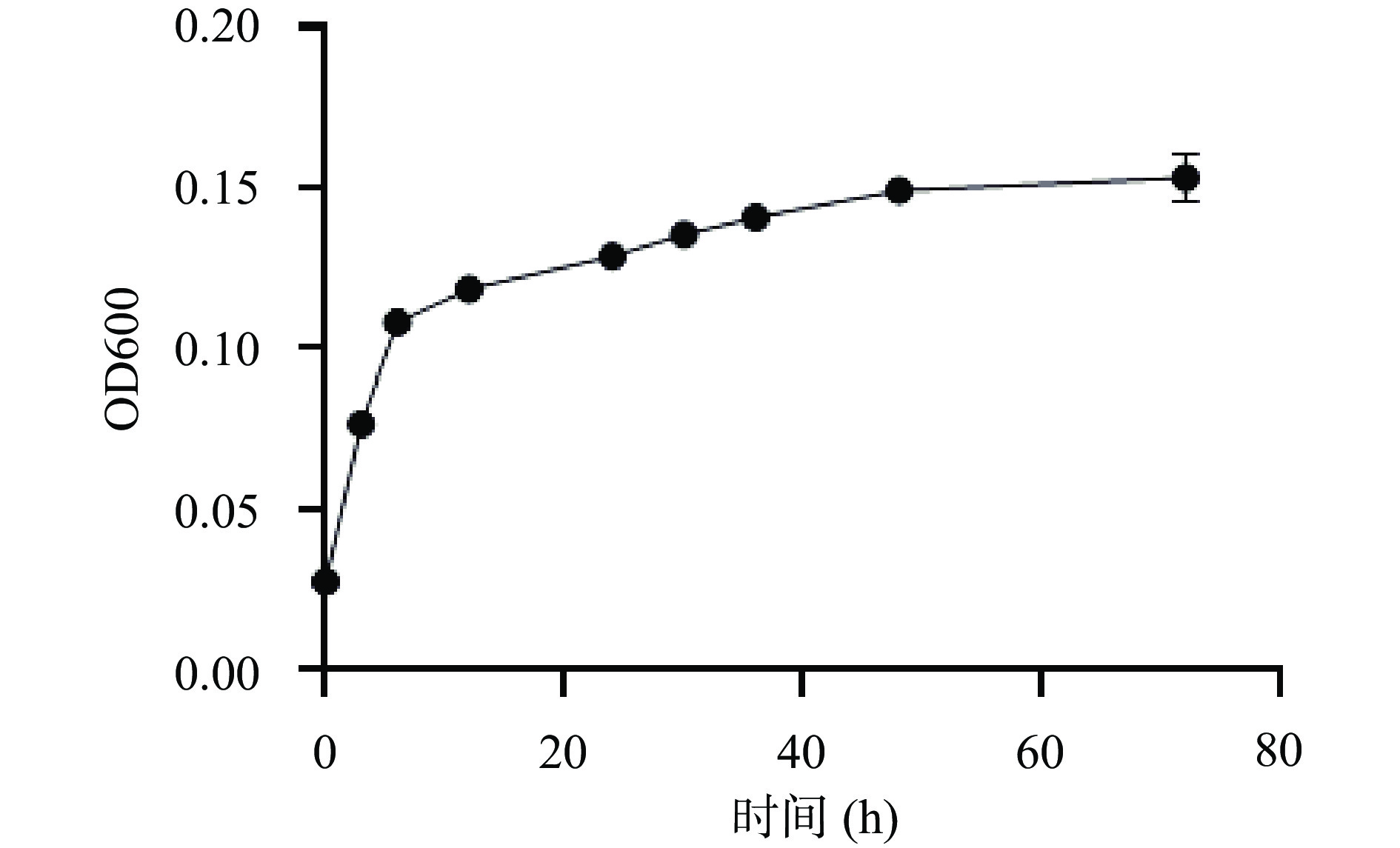
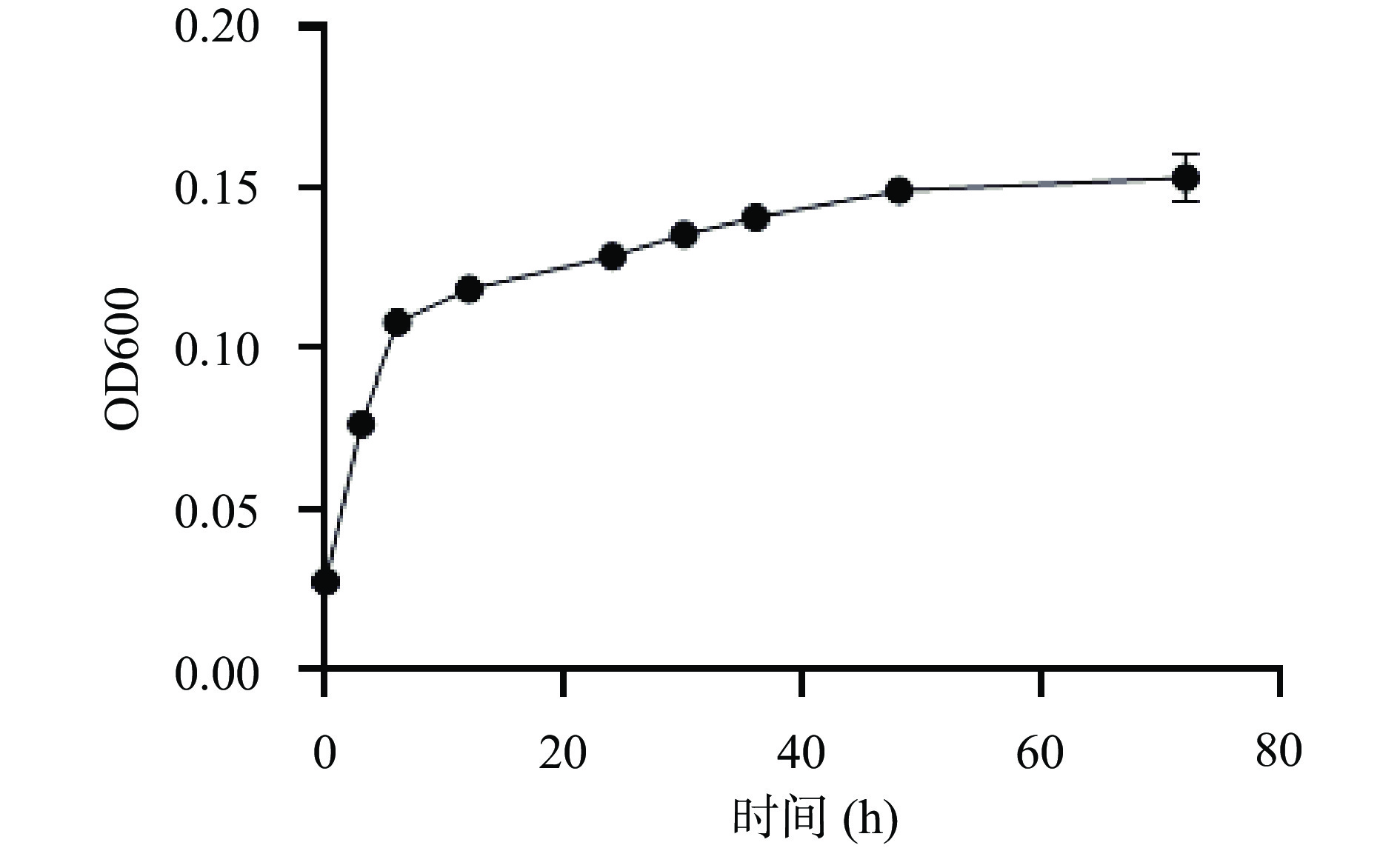
OD600可以用来描述霍氏肠杆菌4-2-1在含有枸杞多糖的培养基中的生长特性[25]。而从图5可以发现,霍氏肠杆菌4-2-1在36 h后基本进入稳定期,在0~12 h处于对数生长期,达到平台期时菌株的OD600在0.15左右。由此可知,霍氏肠杆菌4-2-1可以在含枸杞多糖的培养基中很好的生长,并且存活率较高,所测结果和岳喜庆等[26]筛选出霍氏肠杆菌的生长趋势相似。
由上可知,细菌总量增长,说明霍氏肠杆菌4-2-1能在含枸杞多糖的唯一碳源培养基中生长。
2.3.2 霍氏肠杆菌4-2-1发酵枸杞多糖过程中pH、多糖含量、还原糖含量的变化
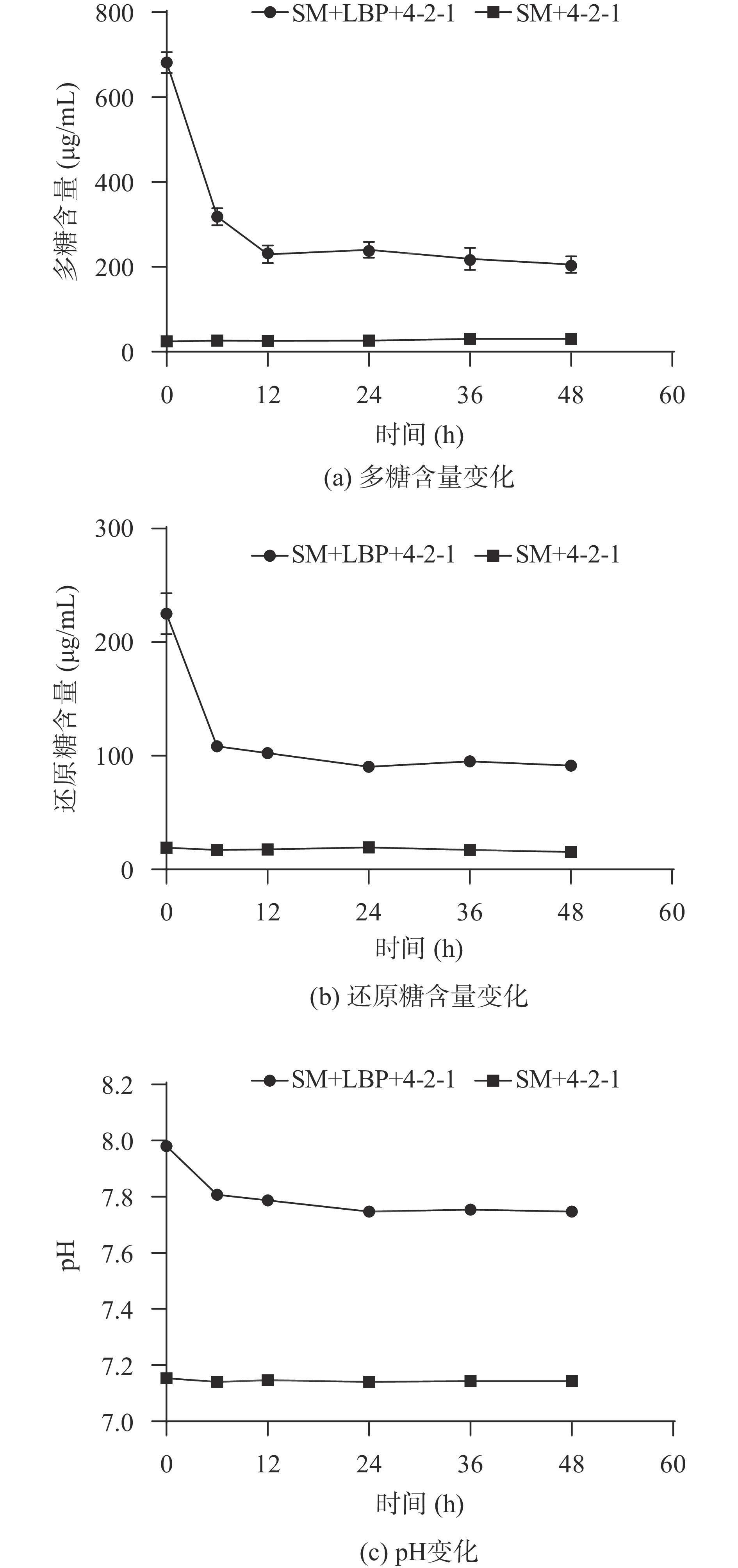
在发酵过程中,检测多糖含量、还原糖含量和pH的动态变化。如图6(a)所示,多糖的初始含量在682 μg/mL左右,此后,在12 h的发酵期间内,多糖含量逐渐降低,逐渐趋于稳定,剩余多糖含量在205.7 μg/mL左右,之前也有研究表明,多糖含量的变化与粪便微生物的作用息息相关[27]。同样的,还原糖也由刚开始的最高含量逐渐降低,直到24 h(图6(b))。通过发酵后多糖和还原糖的分解程度,说明霍氏肠杆菌4-2-1可在枸杞多糖作为唯一碳源的培养基上进行生长。
多糖在肠道里可以被肠道菌选择利用,引起pH的变化[28-29],因此,pH是反应发酵过程的一个重要指标。如图6(c)所示,发酵12 h后,发酵组的pH从7.98降至7.79,在24 h发酵结束时,降低至7.75并保持稳定。然而在整个发酵过程中,对照组的pH保持不变,和之前的荔枝多糖作为益生元的研究相似[30]。表明枸杞多糖可被肠道菌霍氏肠杆菌4-2-1发酵利用,引起培养基相应pH的变化,并可能产生相应的有机酸[31-32]。
2.3.3 霍氏肠杆菌4-2-1发酵枸杞多糖过程中枸杞多糖分子量变化
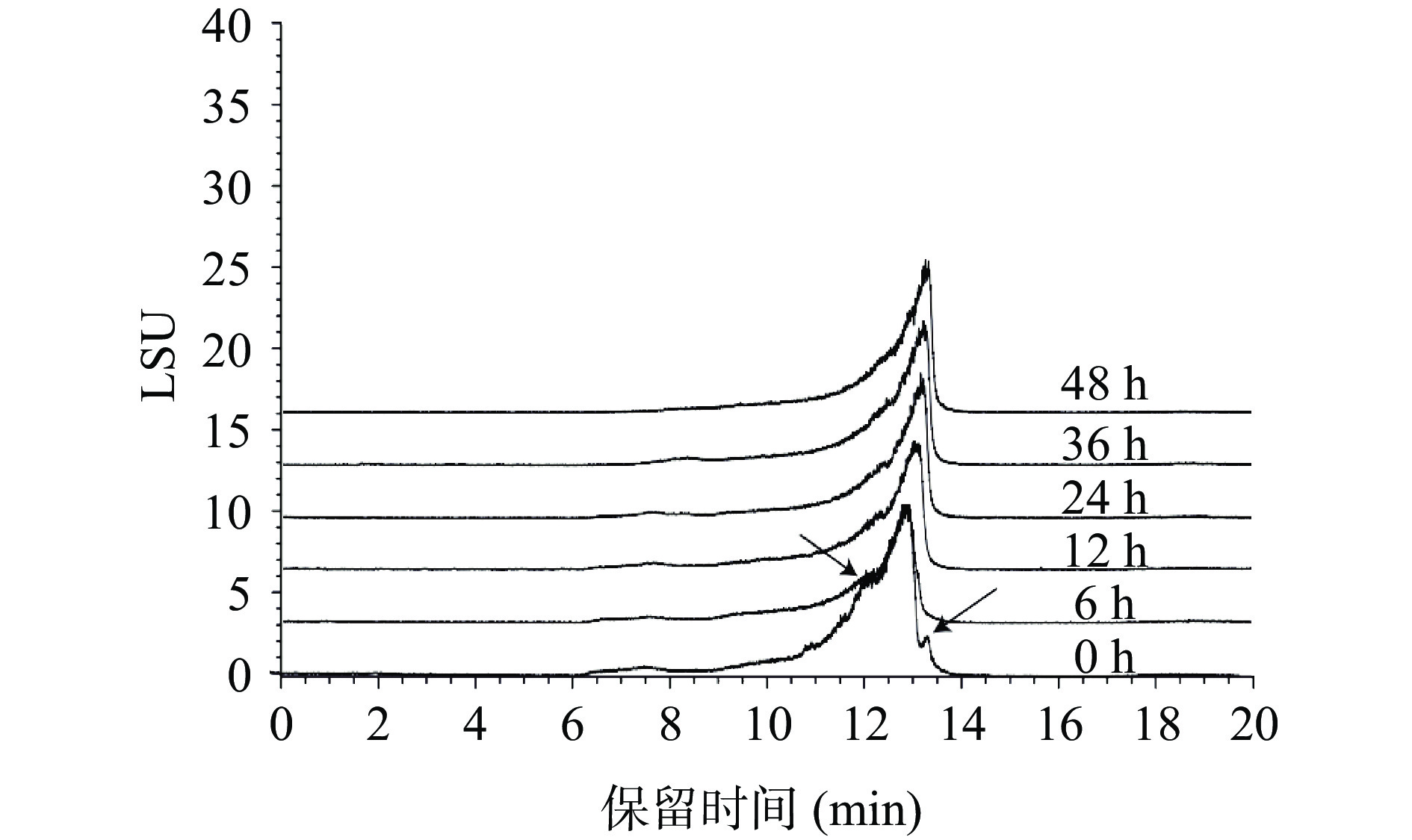
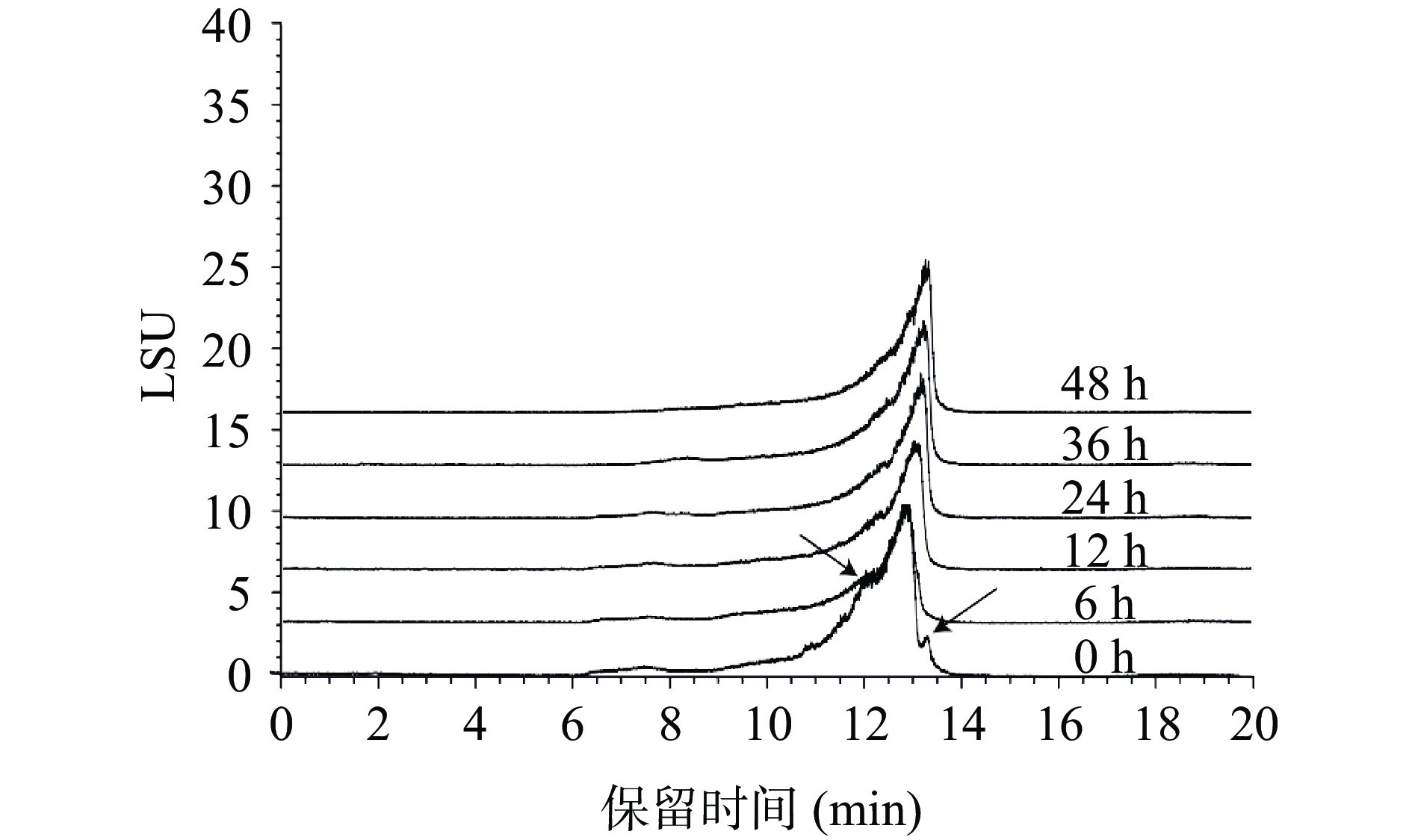
对发酵后枸杞多糖的分子量进行检测,结果如图7。在发酵的过程中,枸杞多糖的含量随着霍氏肠杆菌的发酵时间推移而降低,分子量也从8.02 kDa变为3.44 kDa,表明枸杞多糖在霍氏肠杆菌的发酵下被分解,由此可以推测,霍氏肠杆菌对于枸杞多糖在人体肠道的消化吸收发挥着重要的作用[33]。
![]() 图 7 不同时间点枸杞多糖分子量测定Figure 7. Determination of molecular weight of Lycium barbarum polysaccharide at different time points
图 7 不同时间点枸杞多糖分子量测定Figure 7. Determination of molecular weight of Lycium barbarum polysaccharide at different time points3. 结论
多糖经微生物发酵后可以被人体利用并发挥有益作用。因此研究对枸杞多糖的发酵是了解枸杞多糖在体内作用的重要部分。本研究从人体粪便样品中分离出一株对枸杞多糖具有很好发酵作用的霍氏肠杆菌4-2-1,检测培养基中细菌生长曲线、pH变化、多糖含量、还原糖含量和分子量变化。结果发现,该菌为革兰氏阴性菌霍氏肠杆菌,能在以枸杞多糖为碳源的培养基中进行生长,且随着发酵的进行,细菌数量显著增加,导致发酵体系内pH由7.98变化至7.79,多糖含量、还原糖含量和多糖分子量发生变化,分别降低了30.2%、40.6%和42.9%,说明从人肠道粪便样品中分离的霍氏肠杆菌4-2-1能对枸杞多糖进行发酵。综上,人肠道来源的霍氏肠杆菌4-2-1可发酵枸杞多糖,这可能部分解释了枸杞多糖作为益生元对机体健康的改善作用,为多糖的生理活性进一步提供依据,但详细的机制还需要进一步研究。
-
![]()
图 1 菌株筛选过程中多糖含量的变化
Figure 1. Changes in polysaccharide content during strain screening
![]()
图 3 1%琼脂凝胶电泳图
注:M:2000 DNAMaker;1:所提菌株基因组。
Figure 3. Agarose gel electrophoresis diadram
![]()
图 4 菌株4-2-1的16S rRNA基因序列系统发育树
Figure 4. Phylogenetic tree of Enterobacter 16S rRNA gene sequence of strain 4-2-1
-
[1] YAO, R, HEINRICI M, WECKERLE C S. The genus Lycium as food and medicine: A botanical, ethnobotanical and historical review[J]. J Ethnopharmacol,2018,212:50−66. doi: 10.1016/j.jep.2017.10.010
[2] QIAN, D, ZHAO Y X, YANG G, et al. Systematic review of chemical constituents in the genus Lycium (Solanaceae)[J]. Molecules,2017,22(6):911−33. doi: 10.3390/molecules22060911
[3] LIU, L, SHA X Y, WU Y N, et al. Lycium barbarum polysaccharides protects retinal ganglion cells against oxidative stress injury[J]. Neural Regen Res,2020,15(8):1526−1531. doi: 10.4103/1673-5374.274349
[4] 孙青, 陈亮, 陈莎莎. 枸杞多糖通过核因子κB信号通路对过氧化氢诱导的神经细胞损伤保护作用的研究[J]. 中国糖尿病杂志,2021,29(9):692−699. [SUN Q, CHEN L, CHEN S S. Protective effect of Lycium barbarum polysaccharide on H2O2 induced neuronal injury via NF-κB signaling pathway[J]. Department of Pharmacy,2021,29(9):692−699. doi: 10.3969/j.issn.1006-6187.2021.09.009 [5] GAN, F, LIU Q, LIU Y H, et al. Lycium barbarum polysaccharides improve CCl4-induced liver fibrosis, inflammatory response and TLRs/NF-kB signaling pathway expression in wistar rats[J]. Life Sci,2018,192:205−212. doi: 10.1016/j.lfs.2017.11.047
[6] DING Y, YAN Y M, CHEN D, et al. Modulating effects of polysaccharides from the fruits of Lycium barbarum on the immune response and gut microbiota in cyclophosphamide-treated mice[J]. Food Funct,2019,10(6):3671−3683. doi: 10.1039/C9FO00638A
[7] GAO L L, MA J M, FAN Y M, et al. Lycium barbarum polysaccharide combined with aerobic exercise ameliorated nonalcoholic fatty liver disease through restoring gut microbiota, intestinal barrier and inhibiting hepatic inflammation[J]. Int J Biol Macromol,2021,183:1379−1392. doi: 10.1016/j.ijbiomac.2021.05.066
[8] MASCI A. CARRADORI S, CASADEI M A, et al. Lycium barbarum polysaccharides: Extraction, purification, structural characterisation and evidence about hypoglycaemic and hypolipidaemic effects. A review[J]. Food Chem,2018,254:377−389. doi: 10.1016/j.foodchem.2018.01.176
[9] ZHAO F, GUAN S Z, FU Y J, et al. Lycium barbarum polysaccharide attenuates emotional injury of offspring elicited by prenatal chronic stress in rats via regulation of gut microbiota[J]. Biomed Pharmacother,2021,143:112087. doi: 10.1016/j.biopha.2021.112087
[10] GUERGOLETTO K B, COSTABILE A, FLORES G, et al. In vitro fermentation of jucara pulp (Euterpe edulis) by human colonic microbiota[J]. Food Chem,2016,196:251−258. doi: 10.1016/j.foodchem.2015.09.048
[11] 汪芳芳. 霍氏肠杆菌降解叶黄素的研究[D]. 郑州: 郑州轻工业学院, 2017. WANG F F. Study on degradation of lutein by Enterobacte[D]. Zhengzhou: Zhengzhou University of Light Industry, 2017.
[12] 岳喜庆, 王桐, 单提波, 等, 梁国印. 霍氏肠杆菌WM1产青霉素酶发酵条件的研究[J]. 中国酿造,2010(8):91−93. [YUE X Q, WANG T, SHAN T B, et al. Fermentation conditions of penicillinase by Acinetobacter calcoaceticus WM1[J]. Brewed in China,2010(8):91−93. doi: 10.3969/j.issn.0254-5071.2010.08.027 [13] LANKELMA J M, NIEUWDORP M, DE VOS W M, et al. The gut microbiota in internal medicine: Implications for health and disease[J]. Neth J Med,2015,73(2):61−8.
[14] LOVEGROVE A, EDWARDS, C H, NONI I D. et al. Role of polysaccharides in food, digestion, and health[J]. Crit Rev Food Sci Nutr,2017,57(2):237−253. doi: 10.1080/10408398.2014.939263
[15] RAO T P, QUARTARONE G, Role of guar fiber in improving digestive health and function[J]. Nutrition, 2019, 59: 158−169.
[16] FU X, CAO C L, REN B B, et al. Structural characterization andin vitro fermentation of a novel polysaccharide from Sargassum thunbergii and its impact on gut microbiota[J]. Carbohydr Polym,2018,183:230−239. doi: 10.1016/j.carbpol.2017.12.048
[17] ZHOU X, ZHANG Z, HUANG F H, et al. In vitro digestion and fermentation by human fecal microbiota of polysaccharides from flaxseed[J]. Molecules, 2020, 25(19).
[18] LUO Q, CAI Y, YAN J, et al. Hypoglycemic and hypolipidemic effects and antioxidant activity of fruit extracts from Lycium barbarum[J]. Life Sci,2004,76(2):137−49. doi: 10.1016/j.lfs.2004.04.056
[19] DING Y, YAN Y M, PENG Y J, et al. In vitro digestion under simulated saliva, gastric and small intestinal conditions and fermentation by human gut microbiota of polysaccharides from the fruits of Lycium barbarum[J]. Int J Biol Macromol,2019,125:751−760. doi: 10.1016/j.ijbiomac.2018.12.081
[20] 东秀珠, 蔡妙英. 常见细菌系统鉴定手册[M] . 北京: 科学出版社, 2001: 66 - 95. DONG X Z, CAI M Y. Handbook of systematic identification of common bacteria [M]. Beijing: Science Press, 2001: 66−95.
[21] R E 布坎南. 伯杰细菌鉴定手册[M]. 北京: 科学出版社, 1984: 450-454. R E BU K N. Berger of berger handbook of bacterial identification [M]. Beijing: Science Press, 1984: 450−454.
[22] MASUKO T, MINAMI A, IWASAKI N, et al. Carbohydrate analysis by a phenol-sulfuric acid method in microplate format[J]. Anal Biochem,2005,339(1):69−72. doi: 10.1016/j.ab.2004.12.001
[23] 牟佳红, 梁安雯, 覃超琳, 等. 酶解与发酵联合处理对黑木耳还原糖含量及抗氧化性能的影响[J]. 食品工业科技, 2022, 43(7): 139−147. MOU J H, LIANG AN W, QIN C L, et al. Study on the effect of enzymatic hydrolysis combined with fermentation treatment on reducing sugar content and antioxidant performance of Auricularia auricula[J]. Science and Technology of Food Industry, 2022, 43(7): 139−147.
[24] 唐雨薇. 黄芪均一多糖分离鉴定及微生态调节作用研究[D]. 佳木斯: 佳木斯大学, 2014. TANG Y W. Researching on isolation, identification and micro-ecological regulation of homogeneius polysacccharides from Astragalus[D]. Jiamusi: Jiamusi University, 2014.
[25] RöSCH C, TAVERNE N, VENEMA K, et al. Effects of in vitro fermentation of barley β-glucan and sugar beet pectin using human fecal inocula on cytokine expression by dendritic cells[J]. Molecular Nutrition & Food Research, 2017, 61(1).
[26] 岳喜庆, 王桐, 单提波等. 一株青霉素钠降解菌的分离与鉴定[J]. 食品工业科技,2010,31(6):169−171,175. [YUE X Q, WANG T, SHAN T B, et al. Isolation and identification of a penicillin-degrading bacteria strain[J]. Science and Technology of Food Industry,2010,31(6):169−171,175. doi: 10.13386/j.issn1002-0306.2010.06.094 [27] VANDEPUTTE D, FALONY G, VIEIRA-SILVA S, et al. Prebiotic inulin-type fructans induce specific changes in the human gut microbiota[J]. Gut,2017,66(11):1968−1974. doi: 10.1136/gutjnl-2016-313271
[28] MAKKL K, DEEHAN E C, WALTER J, et al. The impact of dietary fiber on gut microbiota in host health and disease[J]. Cell Host Microbe,2018,23(6):705−715. doi: 10.1016/j.chom.2018.05.012
[29] ZHENG L X, CHEN X Q, ZHANG Z L. Current trends in marine algae polysaccharides: The digestive tract, microbial catabolism, and prebiotic potential[J]. Int J Biol Macromol,2020,151:344−354. doi: 10.1016/j.ijbiomac.2020.02.168
[30] B GULLÓN, P GULLÓN, TAVARIA F, et al. Structural features and assessment of prebiotic activity of refined arabinoxylooligosaccharides from wheat bran[J]. Journal of Functional Foods,2014,6:438−449. doi: 10.1016/j.jff.2013.11.010
[31] GU F, BOREWICZ K, RICHTER B, et al. In vitro fermentation behavior of isomalto/malto-polysaccharides using human fecal inoculum indicates prebiotic potential[J]. Mol Nutr Food Res,2018,62(12):1800232. doi: 10.1002/mnfr.201800232
[32] HUANG F, HONG R Y, YI Y, et al. In vitro digestion and human gut microbiota fermentation of longan pulp polysaccharides as affected by Lactobacillus fermentum fermentation[J]. Int J Biol Macromol,2020,147:363−368. doi: 10.1016/j.ijbiomac.2020.01.059
[33] ZHU W, ZHOU S X, LIU J H, et al. Prebiotic, immuno-stimulating and gut microbiota-modulating effects of Lycium barbarum polysaccharide[J]. Biomed Pharmacother,2020,121:109591. doi: 10.1016/j.biopha.2019.109591
 下载:
下载:


 下载:
下载:

计量
- 文章访问数: 237
- HTML全文浏览量: 124
- PDF下载量: 20
