


Study on Immune-enhancing Activity of Gonad Saponins from Female of Apostichopus japonicus
-
摘要: 目的:以仿刺参雌性生殖腺皂苷(gonad saponins from female of Apostichopus japonicus,AGS)为研究对象,探究其免疫增强活性。方法:通过对巨噬细胞吞噬能力和细胞因子的测定,研究AGS的体外免疫调节活性;通过对免疫低下小鼠胸腺、脾脏指数,碳廓清指数与吞噬指数,腹腔巨噬细胞增殖活性,脾细胞增殖活性,自然杀伤细胞(natural killer cell,NK cell)活力,T淋巴细胞亚群水平及小鼠血清细胞因子的测定,分析AGS在小鼠体内的免疫调节活性。结果:体外实验表明,AGS能提高小鼠体外巨噬细胞吞噬能力,促进TNF-α、IL-6、IFN-γ的分泌,且各组与正常组相比差异显著(P<0.05);体内实验表明,与模型组相比,中剂量组和高剂量组AGS均能显著提高小鼠胸腺、脾脏指数、碳廓清指数和吞噬指数(P<0.05),显著增强腹腔巨噬细胞和脾细胞增殖活性(P<0.05),显著增强NK细胞的活性(P<0.05),显著提高CD4+/CD8+水平(P<0.05),显著促进TNF-α、IL-6、IFN-γ的分泌(P<0.05)。结论:AGS在体外和体内都表现出增强免疫的活性。Abstract: Objective: Taking gonad saponins from female of Apostichopus japonicus (AGS) as the research object to explore its immune-enhancing activity. Methods: The immunoregulatory activity of AGS in vitro was studied by measuring the phagocytosis and cytokines of macrophages. Thereafter the immunoregulatory activity of AGS in immunocompromised mice was analyzed by measuring thymus and spleen index, carbon clearance index and phagocytic index, proliferation activity of peritoneal macrophages, spleen cells, NK cell activity, T lymphocyte subsets level and serum cytokines of mice. Results: In vitro experiments showed that AGS could improve the phagocytic ability of macrophages and enhance the secretion of TNF-α, IL-6 and IFN-γ, and each group was significantly different from the normal group (P<0.05). In vivo experiments showed that compared with model group, AGS in middle dose group and high dose group could significantly increase thymus and spleen index, carbon clearance index and phagocytosis index (P<0.05), significantly enhance the proliferation activity of peritoneal macrophages and spleen cells (P<0.05), significantly enhance the activity of NK cells (P<0.05), significantly increase the level of CD4+/CD8+ (P<0.05), and significantly promote the secretion of TNF-α, IL-6, and IFN-γ (P<0.05). Conclusion: AGS exhibits immune-enhancing activity both in vitro and in vivo.
-
Keywords:
- Apostichopus japonicus /
- gonad /
- saponin /
- immune-enhancing activity
-
仿刺参(Apostichopus japonicus)隶属于棘皮动物门,海参纲,楯手目,刺参科[1],是我国北方重要的经济产物,含有多种生物活性成分,包括类胡萝卜素、多糖、多肽、脂肪酸、皂苷等[2]。其中仿刺参皂苷具有抗肿瘤[3]、抗菌[4]等生物活性,而关于免疫增强活性的研究相对缺乏。生殖腺是海参加工过程中产生的副产物,其来源丰富但相关开发利用主要集中于多肽和多糖的制备、分离及相关活性[5-6],而关于生殖腺皂苷的报道研究非常少。
免疫低下由参与机体免疫的免疫器官、免疫细胞、免疫因子等受到损伤,导致其识别和排除异物、抵御病原体的能力下降所致[7]。活性物质作为免疫调节剂参与机体免疫,是增强机体抗病能力的有效途径[8]。人参皂苷、竹节参皂苷、革皮氏海参皂苷等都已被证实具有调节免疫的作用[9-10],免疫调节主要通过调节免疫器官、免疫细胞和免疫分子等方面发挥作用[11]。而仿刺参生殖腺皂苷在免疫调节方面的作用和机制还鲜有报道。
本论文通过测定仿刺参雌性生殖腺皂苷(gonad saponins from female of Apostichopus japonicus,AGS)对巨噬细胞吞噬活性、巨噬细胞细胞因子TNF-α、IL-6、IFN-γ分泌的影响,探究体外免疫增强活性;通过建立小鼠免疫低下模型,研究其对免疫低下小鼠胸腺、脾脏指数,碳廓清指数与吞噬指数,腹腔巨噬细胞吞噬能力和增殖活性,脾细胞增殖活性,自然杀伤细胞活力,T淋巴细胞亚群水平,小鼠血清细胞因子TNF-α、IL-6、IFN-γ等指标的影响,探究其对免疫低下小鼠免疫能力的作用。通过体内和体外免疫增强活性的研究,以期为其免疫调节相关活性研究提供参考依据。
1. 材料与方法
1.1 材料与仪器
巨噬细胞RAW264.7、人白血病细胞K562、NK细胞 上海雅吉生物科技有限公司;18~20 g SPF级雌性昆明小鼠 济南朋悦实验动物繁育有限公司[许可证号:SCXK(鲁)20190003];注射用环磷酰胺 德国Baxter Oncology公司;FITC anti-mouse CD3 Antibody、PE anti-mouse CD4 Antibody、APC anti-mouse CD8a Antibody 美国Biolegend公司;Cell Counting Kit-8(CCK-8)试剂盒 美国Glpbio公司;RPMI-1640培养基 美国Sigma-Aldrich公司;磷酸盐缓冲溶液 武汉博士德生物工程有限公司;胎牛血清 北京全式金生物技术有限公司;印度墨汁 生工生物工程(上海)股份有限公司;红细胞裂解液、脂多糖(Lipopolysaccharide,LPS)、青链霉素混合液、小鼠肿瘤坏死因子α ELASA试剂盒、小鼠白细胞介素6 ELISA试剂盒、小鼠干扰素γ ELISA试剂盒 北京索莱宝科技有限公司。
ME204E型电子天平 梅特勒-托利多仪器(上海)有限公司;1510型酶标仪、HERACELL 150i型二氧化碳培养箱 赛默飞世尔(中国)有限公司;FC500型流式细胞仪 美国贝克曼库尔特公司;5430R型离心机 德国Eppendorf公司。
1.2 实验方法
1.2.1 AGS的制备
本实验用AGS由实验室前期制备:60%乙醇冷浸提取仿刺参雌性生殖腺冻干粉→旋蒸至浸膏状→分散于水中→水饱和正丁醇萃取→冻干正丁醇层→制得AGS,为淡黄色粉末,Molish反应出现紫色环。
1.2.2 巨噬细胞RAW264.7体外中性红吞噬实验
参考翟星辰[12]的方法,利用RPMI-1640完全培养基调整细胞浓度为5×104个/mL并接种于96孔板中,于37 ℃、5% CO2条件下贴壁培养12 h。设置含有不同终浓度样品组(200、500、1000 μg/mL)、正常组(培养基)和LPS(1 μg/mL)阳性对照组,继续培养48 h,吸去上清液,并于每孔中加入0.1%中性红溶液。在培养箱内孵育3 h后,吸去上清并用PBS清洗细胞2次。向每孔中加入细胞裂解液1 mL,细胞裂解液为乙酸:乙醇=1:1(v/v)。将96孔板置于4 ℃冰箱内,12 h后于540 nm处测定吸光度值。
1.2.3 巨噬细胞RAW264.7体外TNF-α,IL-6,IFN-γ测定
设置含有不同终浓度(200、500、1000 μg/mL)AGS、正常组(培养基)和LPS(1 μg/mL)阳性对照组,根据小鼠肿瘤坏死因子α ELASA试剂盒、小鼠白细胞介素6 ELISA试剂盒、小鼠干扰素γ ELISA试剂盒分别测定体外继续培养48 h后RAW 264.7细胞的TNF-α,IL-6,IFN-γ的分泌。
1.2.4 动物模型建立
将雌性昆明小鼠分为5组,正常组,模型对照组,AGS低、中、高剂量组(0.5,5,20 mg/kg),每组12只小鼠。本实验遵守动物福利3R原则、符合国际实验动物伦理学要求,并由烟台市百吉林生命科学研究院实验动物伦理委员会审查批准。小鼠在空调动物饲养室中同室群笼饲养,自由饮水。每天定时通风,室内温度保持在22~24 ℃。湿度45%~60%。保持12 h黑暗/12 h照明的昼夜节律(8:00~20:00亮灯)。小鼠适应性饲养7 d。在第8~10 d,空白组小鼠腹腔注射给予生理盐水,其余组小鼠腹腔内注射80 mg/kg/d的环磷酰胺,建立环磷酰胺诱导的免疫低下模型。在第11~20 d,进行给药处理,处理方法如表1所示。
表 1 各组小鼠给药处理表Table 1. Dosing and treatment of mice in each group序号 组别 是否注射环磷酰胺 第11~20 d灌胃 1 正常组 否 去离子水 2 模型对照组 是 去离子水 3 低剂量组 是 0.5 mg/kg 4 中剂量组 是 5 mg/kg 5 高剂量组 是 20 mg/kg 1.2.5 胸腺、脾脏指数的测定
脏器指数的测定参考董婧媛[13]的方法,在最后一次给药后12 h,小鼠禁食脱水,脱臼处死后取出小鼠胸腺和脾脏并称重。按公式1和公式2计算胸腺指数和脾脏指数:
胸腺指数(mg/10gbw)=胸腺质量小鼠体重 (1) 脾脏指数(mg/10gbw)=脾脏质量小鼠体重 (2) 1.2.6 小鼠碳廓清指数与吞噬指数
参考文献[14]的方法,在最后一次给药24 h后,注入浓度为10%的墨汁于小鼠的尾部静脉,墨汁用量为0.1 mL/10 g。提前配制0.1% NaCO3溶液,将2 min和10 min小鼠眼眶取血20 μL分别加入到0.1% NaCO3溶液中并摇匀,于600 nm处测定吸光度值。小鼠脱臼处死后,对肝脏和脾脏进行称重,按公式3和公式4计算碳廓清指数和吞噬指数:
K=lgA1−lgA2t2−t1 (3) α=m1m2+m3×3√K (4) 式中:K—碳廓清指数;A1—2 min时吸光度;A2—10 min时吸光度;t1—2,min;t2—10,min;α—吞噬指数;m1—小鼠体重,g;m2—肝脏重量,g;m3—脾脏重量,g。
1.2.7 小鼠腹腔巨噬细胞增殖活性测定
1.2.7.1 小鼠腹腔巨噬细胞的制备
于75%乙醇中浸泡脱臼处理的小鼠1~2 min,在超净工作台中向小鼠腹腔缓慢注入5 mL预冷的PBS缓冲溶液,轻柔腹部1~2 min,用无菌注射器吸取腹水于离心管中,重复2~3次。在4 ℃、1000 r/min条件下离心5 min,弃上清,重悬细胞于RPMI-1640完全培养基中,置于5% CO2、37 ℃恒温培养箱中培养。
1.2.7.2 巨噬细胞活性的测定
利用RPMI-1640完全培养基调整细胞浓度为2×104个/mL并接种于96孔板中,每孔100 μL。设计空白组和实验组,其中空白组以完全培养基调零,每组设计6个复孔。在5% CO2、37 ℃恒温培养箱中培养48 h后,弃去孔内培养基并向每孔中加入含有10%的CCK-8溶液100 μL,于培养箱中继续培养4 h后利用酶标仪检测450 nm处吸光度[13]。
1.2.8 小鼠脾细胞悬液的制备及增殖测定
1.2.8.1 小鼠脾细胞的制备
于75%乙醇中浸泡脱臼处理的小鼠1~2 min,无菌条件下取出脾脏,过200目细胞筛,置于盛有D-Hank’s液的培养皿中,用D-Hank’s缓冲溶液冲洗细胞筛并收集细胞悬液。在4 ℃、1000 r/min条件下离心5 min,弃上清。在室温条件下,加入5 mL红细胞裂解液裂解3 min,加入1 mL胎牛血清后于4 ℃、1000 r/min条件下离心5 min,重复2~3次。利用RPMI-1640完全培养基重悬细胞,置于5% CO2、37 ℃恒温培养箱中培养。
1.2.8.2 脾细胞活性的测定
利用RPMI-1640完全培养基调整细胞浓度为2×104个/mL并接种于96孔板中,每孔100 μL。设计空白组和实验组,其中空白组以完全培养基调零,每组设计6个复孔。在5% CO2、37 ℃恒温培养箱中培养48 h后,弃去孔内培养基并向每孔中加入含有10%的CCK-8溶液100 μL,于培养箱中继续培养4 h后利用酶标仪检测450 nm处吸光度[13]。
1.2.9 小鼠脾细胞中自然杀伤细胞(NK细胞)活力测定
参考翟星辰[12]的方法,按照脾细胞:人白血病细胞K562=50:1接种于96孔板中,即每孔接种5×105个脾细胞后再接种1×104个人白血病细胞K562于同样的孔板中,两种细胞混合的孔为样品孔,对照孔为各细胞单独培养,培养时间为24 h。按照公式5计算小鼠脾细胞中NK细胞活力:
NK(%)=T−(S−E)T×100 (5) 式中:T—人白血病细胞K562孔吸光度;S—样品孔吸光度;E—脾细胞孔吸光度。
1.2.10 T 淋巴细胞亚群的测定
参考刘仁杰等[15]的方法,按照1.2.8.1的方法制备小鼠脾淋巴细胞单细胞悬液后,将细胞浓度调整为4×107个/mL,加入10 μL FITC anti-mouse CD3、PE anti-mouse CD4、APC anti-mouse CD8a抗体,充分混匀并于4 ℃条件下避光孵育1 h。而后加入1 mL FACS buffer,在4 ℃、2000 r/min条件下离心5 min,收集沉淀。缓慢加入150 mL FACS buffer重悬细胞沉淀,用流式细胞仪检测并计算CD4+、CD8+T淋巴细胞比例。
1.2.11 小鼠血清中TNF-α,IL-6,IFN-γ的测定
将小鼠处死后,取眼球全血3 mL并分离出血清。用ELISA试剂盒检测血清中TNF-α,IL-6,IFN-γ的分泌。
1.3 数据处理
采用Excel 2010整理实验数据,使用SPSS 20.0软件作统计分析,分析方法采用t检验,P<0.05时代表有显著性差异,P<0.01时代表有极显著性差异。使用Origin 2019绘制图形。实验重复六次。
2. 结果与分析
2.1 AGS对巨噬细胞RAW264.7吞噬能力的影响
中性红是一种能被巨噬细胞吞噬的大分子荧光试剂,在540 nm波长下有强吸收峰[12]。中性红实验能反应吞噬细胞的吞噬能力,其吞噬能力与吸光度成正比,巨噬细胞的吞噬能力如表2所示。与未进行处理(正常组)的巨噬细胞RAW264.7相比,AGS各浓度组及LPS刺激的巨噬细胞的吞噬能力都显著增强(P<0.05),且样品组与LPS组无显著差异,说明AGS具有促进巨噬细胞吞噬大分子的能力,与绞股蓝总皂苷增强巨噬细胞吞噬中性红的能力相似[16]。
表 2 AGS对RAW264.7吞噬能力的影响Table 2. Effects of AGS on the phagocytosis of RAW264.7 cells组别 吸光度 正常组 0.282±0.014 200(μg/mL) 0.329±0.018* 500(μg/mL) 0.321±0.006* 1000(μg/mL) 0.320±0.007* LPS 0.338±0.015* 注:*表示与正常组相比,差异显著(P<0.05)。 2.2 AGS对巨噬细胞RAW264.7细胞因子的影响
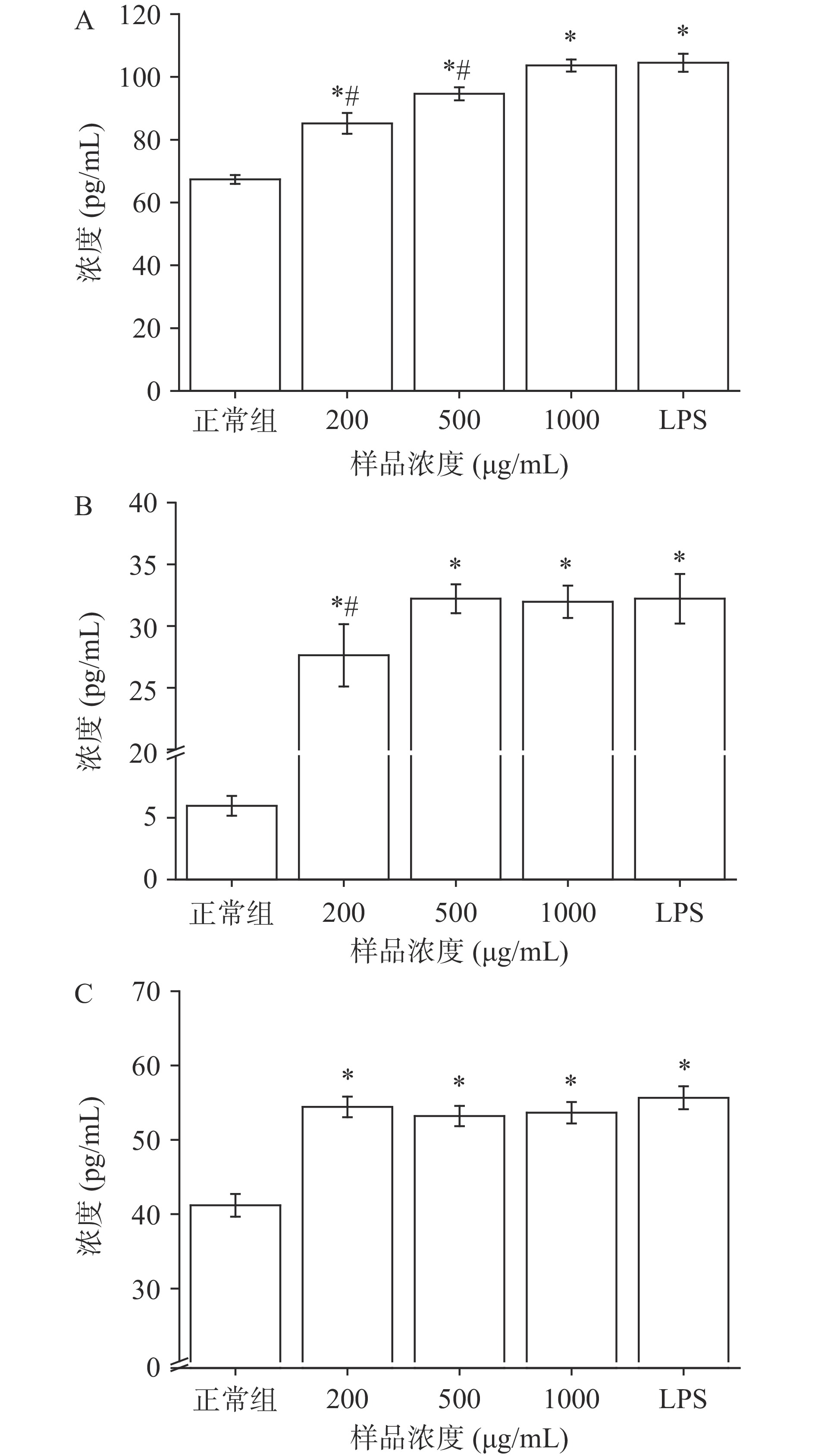
细胞因子主要由免疫细胞和个别非免疫细胞产生,是一类小分子蛋白[17],能与相应受体结合参与细胞间的信息传递,从而形成细胞动态网络,调节细胞的生长和代谢、机体的免疫应答[18]。细胞因子包括肿瘤坏死因子、白细胞介素、干扰素等,能介导炎症反应和天然免疫等。AGS对巨噬细胞RAW264.7细胞因子TNF-α、IL-6、IFN-γ分泌的影响如图1所示。
![]()
TNF-α主要由M1型巨噬细胞产生,在诱导细胞凋亡、与病原体和肿瘤细胞直接作用、清除异常细胞等都发挥了重要功能,能促进下游细胞因子IL-6、IL-1β、黏附因子等的表达[12]。相对于未处理RAW264.7,AGS和LPS组能显著提高TNF-α的含量(P<0.05),且AGS 1000 μg/mL组与LPS组效果无明显差异(P>0.05),说明AGS能促进TNF-α的分泌,与Aminin等[19]研究发现海参单体皂苷具有增强TNF-α分泌的作用相似。
IL-6可由活化的巨噬细胞、T淋巴细胞等产生,能参与和刺激免疫应答的细胞增殖、分化并提高免疫功能,具有诱导B细胞分化、诱导IL-2和IL-2受体表达、增强NK细胞活性、诱导单核细胞分化等重要作用[20]。相对于未处理RAW264.7,AGS和LPS组能显著提高IL-6的含量(P<0.05),且AGS 500和1000 μg/mL组与LPS组效果无明显差异(P>0.05),说明AGS能较好地促进IL-6的分泌,与羊肚菌多糖增强IL-6的作用相似[21]。
干扰素可激活巨噬细胞,增强巨噬细胞的活性,而活化的巨噬细胞又可产生干扰素。IFN-γ可由活化的T细胞和NK细胞产生,除成熟的红细胞以外,IFN-γ分布于几乎所有细胞表面,能通过旁邻方式控制细胞的生长。相对于未处理组RAW264.7,AGS和LPS组能显著提高IFN-γ的含量(P<0.05),且AGS各剂量组与LPS组效果无明显差异(P>0.05),说明AGS能较好地促进IFN-γ分泌。Bhardwaj等[22]发现茶皂苷具有增强IFN-γ分泌的作用,与本研究结果一致。
2.3 AGS对小鼠胸腺指数、脾脏指数的影响
环磷酰胺因其具有较好的细胞毒性作用,普遍应用于肿瘤疾病的治疗之中。但与此同时,环磷酰胺也会导致免疫抑制[23]、骨髓抑制[24]、介导氧化还原反应的平衡从而引起氧化应激等反应[25]。胸腺和脾脏是人体进行免疫调节的外周免疫器官,具有抗感染、造血等重要作用[26],其质量和大小受到免疫细胞增殖和分化的影响[27]。胸腺和脾脏的质量随着年龄的增长而逐渐萎缩,且在应激条件下也能加速萎缩。AGS对免疫低下小鼠胸腺、脾脏指数的影响如表3所示。与正常组相比,免疫低下模型组小鼠的胸腺指数和脾脏指数都显著降低(P<0.05),说明环磷酰胺的注射导致了小鼠胸腺、脾脏的萎缩,质量减轻,表明建模成功。与模型组相比,经AGS的灌胃后,AGS各剂量组小鼠的胸腺、脾脏指数都有所提升,其中中、高剂量AGS灌胃的小鼠差异均显著(P<0.05)。由此可推测出AGS能够缓解由环磷酰胺诱导的免疫低下小鼠的胸腺、脾脏萎缩,质量减轻的损害,其结果与积雪草皂苷的作用相似[28]。
表 3 AGS对免疫低下小鼠脏器指数的影响Table 3. Effects of AGS on organ indices in immunocompromised mice2.4 AGS对小鼠单核-巨噬细胞碳廓清指数和吞噬指数的影响
肝脏、脾脏中的巨噬细胞具有清除碳粒的作用,吞噬能力是评价巨噬细胞的关键指标[29,13]。而碳廓清指数和吞噬指数能反映单核-巨噬细胞的吞噬能力。由表4可知,与正常组相比,模型组小鼠的碳廓清指数和吞噬指数都显著降低(P<0.05)。说明环磷酰胺能降低小鼠巨噬细胞的吞噬能力。与模型组相比,AGS中、高剂量小鼠的碳廓清指数和吞噬指数都显著提高(P<0.05),说明AGS具有缓解环磷酰胺诱导的免疫低下小鼠巨噬细胞吞噬能力的损害的作用。Chen等[30]发现大豆皂苷能显著提升巨噬细胞的吞噬能力,本研究结果与之类似。
表 4 AGS对免疫低下小鼠碳廓清指数、吞噬指数的影响Table 4. Effect of AGS on carbon clearance index and phagocytic index in immunocompromised mice组别 碳廓清指数 吞噬指数 正常组 0.0689±0.0019 0.3551±0.0179 模型组 0.0199±0.0015* 0.0993±0.0119* 低剂量组 0.0210±0.0020* 0.1110±0.0116* 中剂量组 0.0339±0.0030*# 0.1888±0.0069*# 高剂量组 0.0409±0.0014*# 0.2489±0.0266*# 2.5 AGS对小鼠巨噬细胞增殖活性的影响
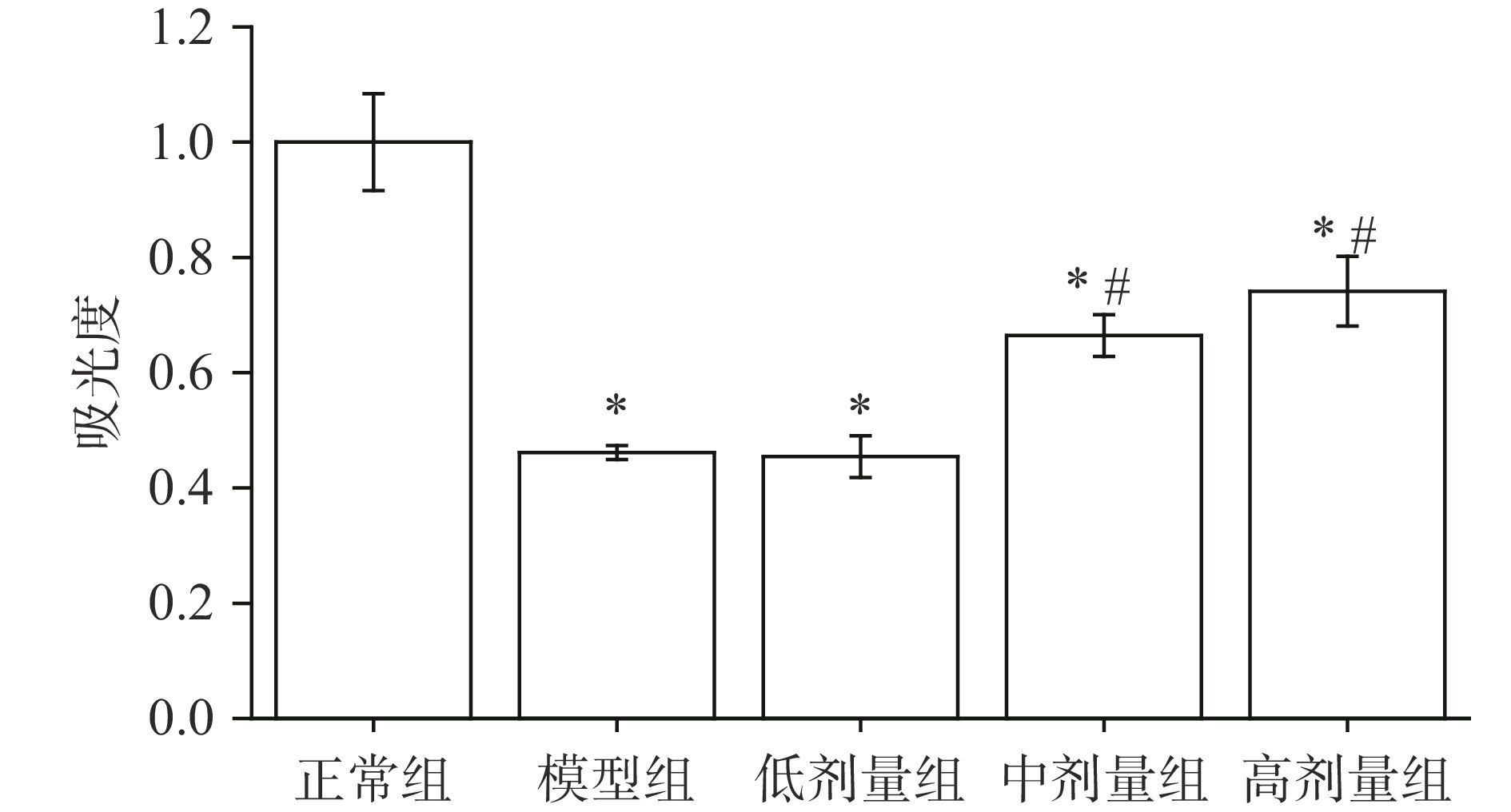
巨噬细胞分布广泛,几乎存在于身体的所有组织中[31],能吞噬和杀死病原体、将抗原呈递给淋巴细胞并释放大量生物活性分子,在特异性免疫和非特异性免疫都起到了重要作用[32]。由图2可知,模型组小组的巨噬细胞增殖活性显著低于正常组小鼠(P<0.05),说明环磷酰胺能降低小鼠巨噬细胞增殖活性。与模型组相比,中、高剂量组小鼠巨噬细胞增殖活性显著升高(P<0.05),说明AGS对注射环磷酰胺小鼠巨噬细胞增殖能力降低具有一定的缓解作用,其结果与海参皂苷对巨噬细胞增殖活性的影响相似[13]。
![]() 图 2 AGS对小鼠巨噬细胞增殖活性的影响Figure 2. Effects of AGS on proliferation of macrophages in mice
图 2 AGS对小鼠巨噬细胞增殖活性的影响Figure 2. Effects of AGS on proliferation of macrophages in mice2.6 AGS对小鼠脾细胞增殖活性的影响
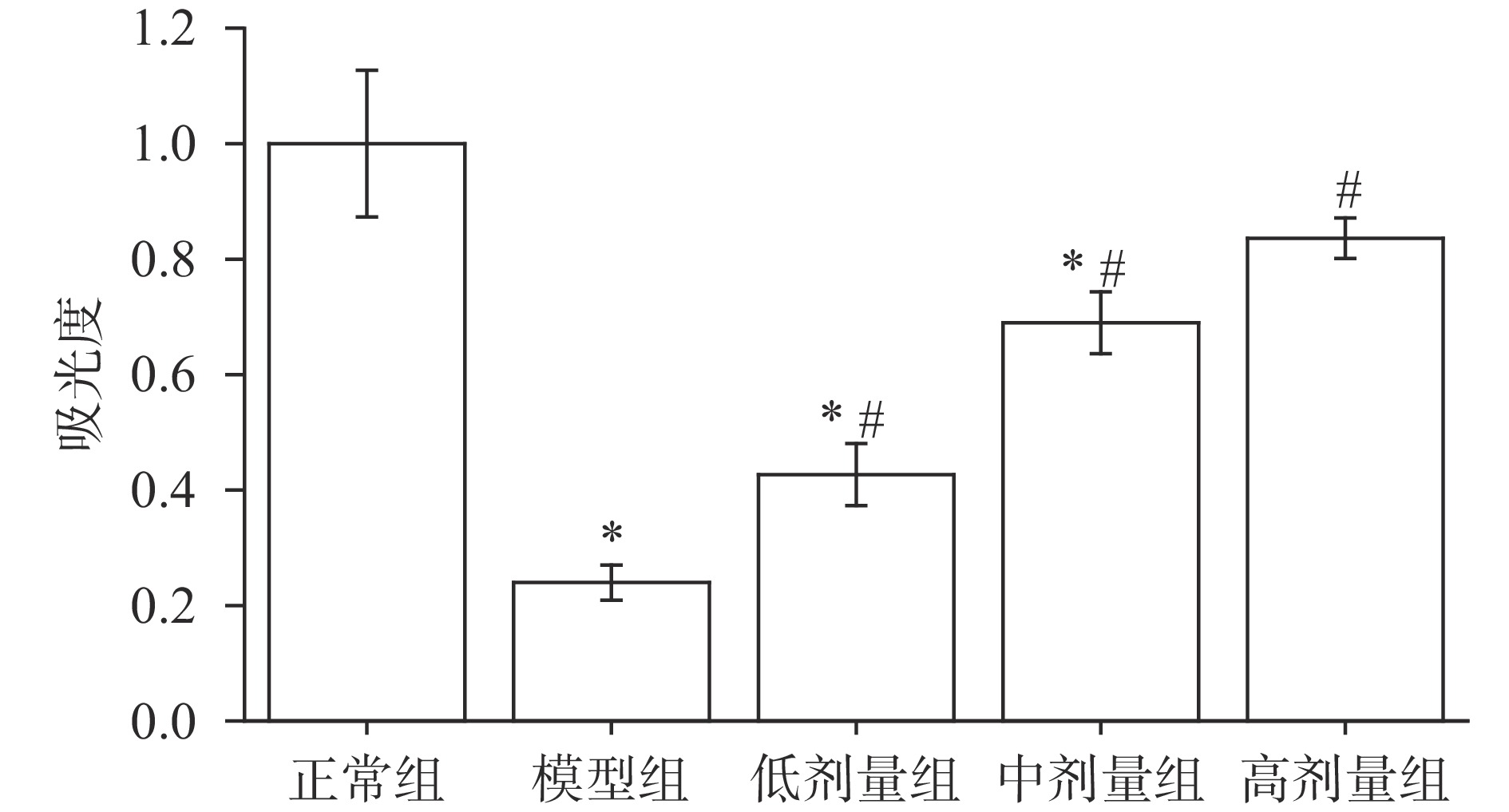
脾细胞中含有多种免疫细胞如巨噬细胞、T淋巴细胞、B淋巴细胞等,对机体的细胞免疫起到重要的调节作用[33]。由图3可知,经环磷酰胺诱导的小鼠脾细胞增殖活性显著低于正常组小鼠(P<0.05)。与模型组相比,低、中、高剂量组小鼠的脾细胞增殖活性都显著提高(P<0.05)。说明AGS能较好地缓解环磷酰胺诱导的小鼠脾细胞增殖活性降低的现象。Rositsa等[34]在研究黄芪皂苷时发现,黄芪皂苷具有增强脾细胞活性的作用,且具有剂量依赖性,本研究结果与之相似。
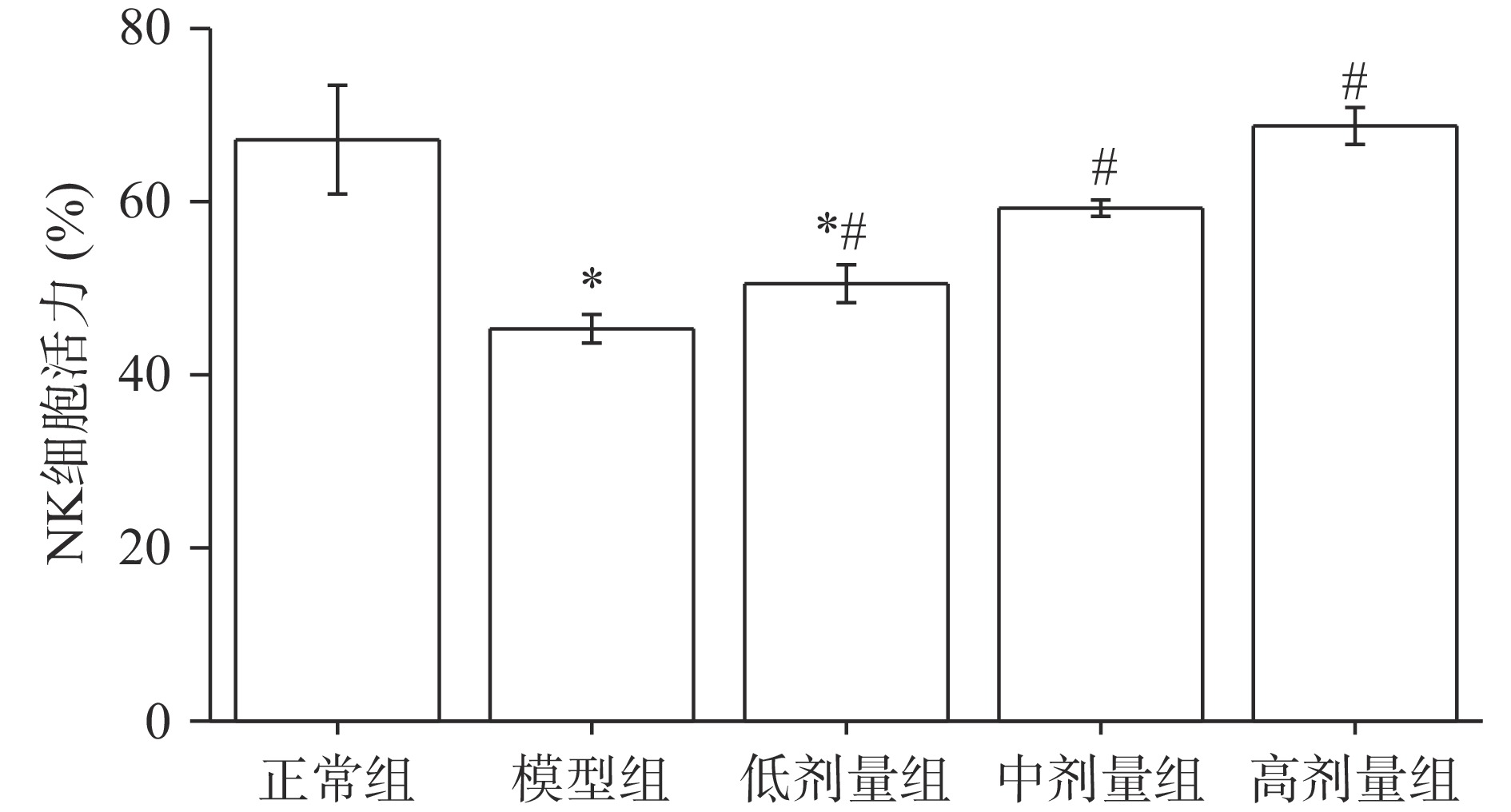
2.7 AGS对小鼠脾细胞中NK细胞活力的影响
NK细胞能对病毒、炎症、肿瘤等靶标进行识别,从而具有抗肿瘤、抗病毒感染等作用[35]。白血病细胞K562是自然杀伤细胞的敏感细胞,NK细胞对其发挥着细胞毒性作用,据此研究NK细胞的活力。由图4可知,与正常组小鼠相比,经环磷酰胺诱导后小鼠NK细胞活力显著降低(P<0.05),说明环磷酰胺能够降低NK细胞的功能。与模型组相比,低、中、高剂量组小鼠的NK细胞活力都有所提高,且差异显著(P<0.05)。另外,经过中、高剂量AGS喂食的小鼠组NK细胞活性与正常组小鼠差异不显著(P>0.05),效果较好。由此可推测,AGS对环磷酰胺诱导的免疫低下小鼠的NK细胞活性降低具有缓解作用,与日本刺参对NK细胞活性增强的研究结果相似[36]。
![]() 图 4 AGS对小鼠脾细胞中自然杀伤细胞(NK细胞)活力的影响Figure 4. Effects of AGS on NK cell activity of splenocytes in mice
图 4 AGS对小鼠脾细胞中自然杀伤细胞(NK细胞)活力的影响Figure 4. Effects of AGS on NK cell activity of splenocytes in mice2.8 AGS对小鼠脾脏T淋巴细胞亚群CD4+,CD8+T细胞的影响
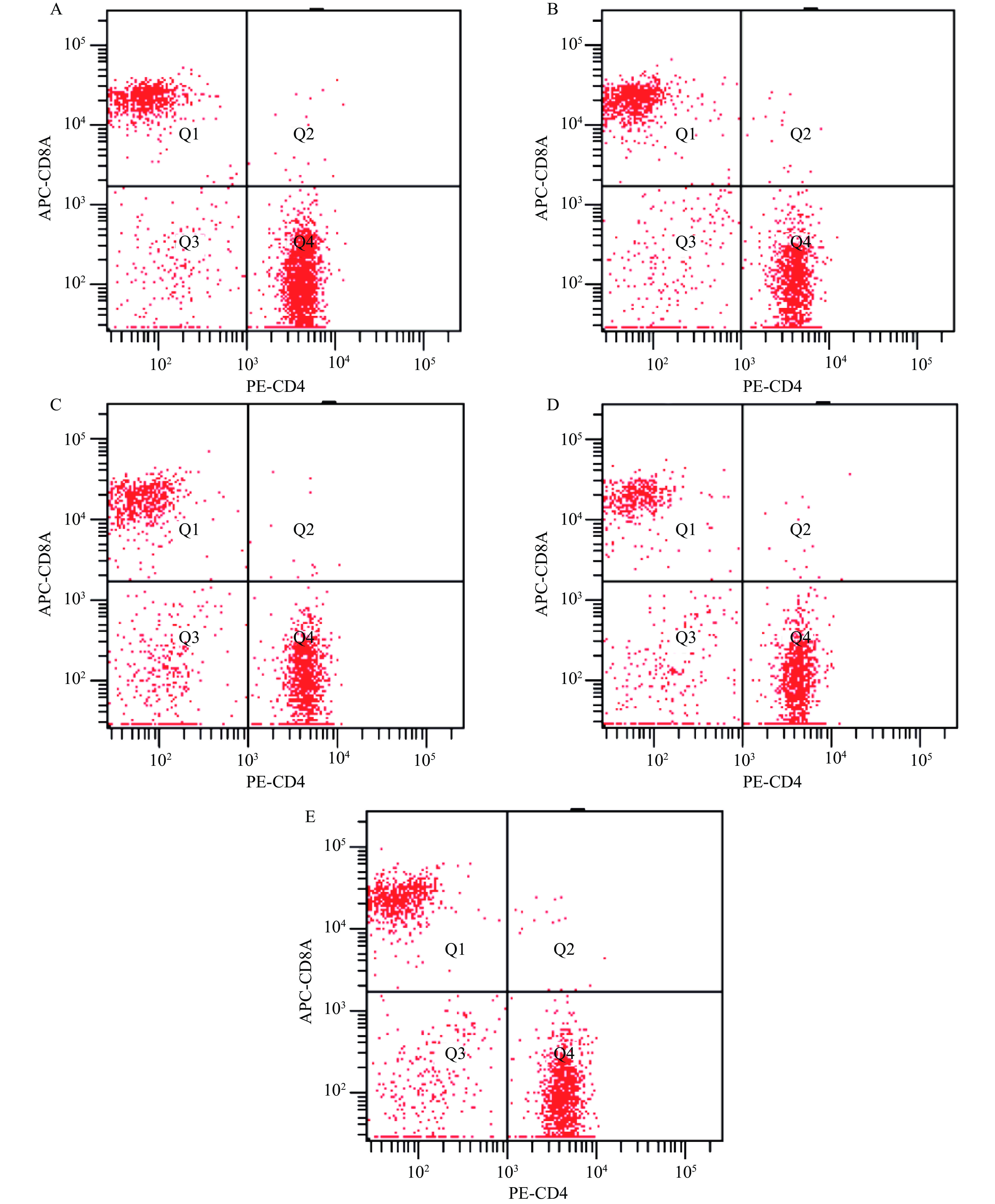
T淋巴细胞参与机体的免疫调节,包含了不同功能的T淋巴细胞亚群。其中,辅助性T细胞和效应T细胞的表面标记抗体为CD4;细胞毒性T细胞和抑制性T细胞的表面标记抗体为CD8[37]。CD4+T细胞和CD8+T细胞相互制约,形成T细胞网络,进而调节机体的细胞免疫的稳定性,且免疫功能的高低能通过CD4+与CD8+的比值反应[38]。经合理设门后,不同处理组小鼠T淋巴细胞亚群分群情况如图5所示。
![]() 图 5 AGS对小鼠脾脏T淋巴细胞亚群的影响注:A:正常组;B:模型组;C:低剂量组;D:中剂量组;E:高剂量组。Figure 5. Effect of AGS on T lymphocyte subsets of spleen in mice
图 5 AGS对小鼠脾脏T淋巴细胞亚群的影响注:A:正常组;B:模型组;C:低剂量组;D:中剂量组;E:高剂量组。Figure 5. Effect of AGS on T lymphocyte subsets of spleen in mice经计算,CD4+、CD8+水平及其比例如表5所示。与正常组相比,模型组小鼠的CD4+水平显著降低(P<0.05);CD8+水平显著升高(P<0.05);CD4+/CD8+水平显著降低(P<0.05),说明环磷酰胺注射的小鼠免疫功能低下。与模型组小鼠相比,AGS低、中、高剂量组小鼠的CD4+水平都有所提高,其中AGS中、高剂量组差异显著(P<0.05),且中剂量组小鼠提升效果最明显;AGS各剂量组CD8+水平均显著降低(P<0.05),其中中剂量组小鼠降低效果最明显;AGS各剂量组CD4+/CD8+水平显著增高(P<0.05),其中中剂量组小鼠升高效果最佳。由此可知,AGS能对环磷酰胺诱导的小鼠免疫低下起到一定的缓解作用,能增强免疫低下小鼠的免疫功能,且中剂量组效果最好。研究表明,皂苷具有一定的毒性[39],高剂量组效果稍差可能的原因是高剂量的AGS表现出轻微的副作用。
表 5 AGS对小鼠脾脏T淋巴细胞亚群的影响Table 5. Effect of AGS on T lymphocyte subsets of spleen in mice组别 CD4+(%) CD8+(%) CD4+/CD8+ 正常组 72.33±0.40 21.63±0.64 3.35±0.11 模型组 56.17±0.57* 35.80±0.26* 1.57±0.02* 低剂量组 58.10±1.21* 27.80±0.87*# 2.09±0.11*# 中剂量组 66.23±0.40*# 22.13±0.57# 2.99±0.09*# 高剂量组 58.43±0.32*# 29.50±0.79*# 1.98±0.04*# 2.9 AGS对小鼠血清细胞因子的影响
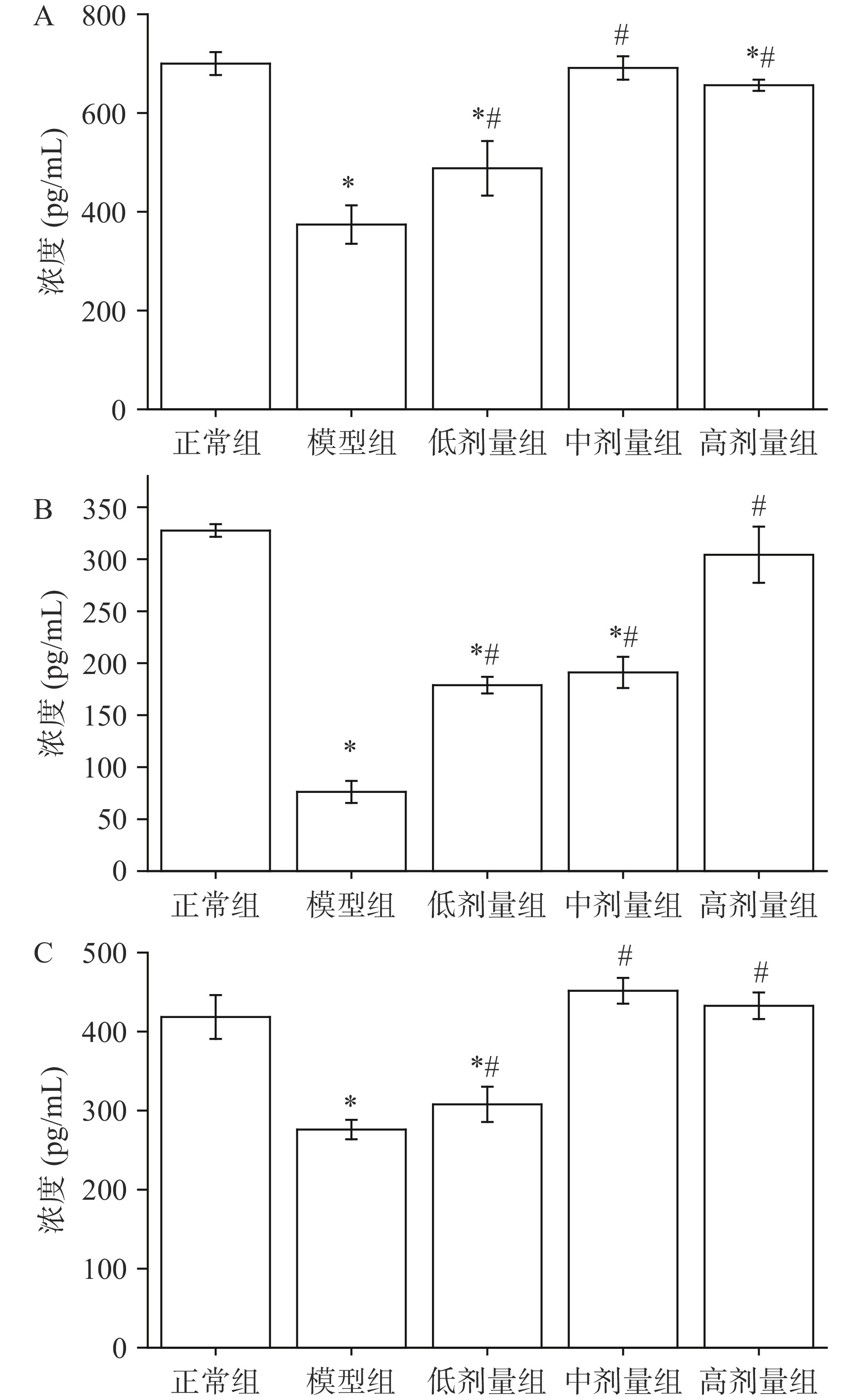
TNF-α、IL-6、IFN-γ在免疫调节、抗肿瘤等多方面都起到重要作用,AGS对小鼠血清的TNF-α、IL-6、IFN-γ分泌影响如图6所示。
![]() 图 6 AGS对小鼠血清细胞因子的影响注:A:TNF-α;B:IL-6;C:IFN-γ;*表示与正常组相比,差异显著(P<0.05);#表示与模型组相比,差异显著(P<0.05)。Figure 6. Effects of AGS the release of cytokines in serum of mice
图 6 AGS对小鼠血清细胞因子的影响注:A:TNF-α;B:IL-6;C:IFN-γ;*表示与正常组相比,差异显著(P<0.05);#表示与模型组相比,差异显著(P<0.05)。Figure 6. Effects of AGS the release of cytokines in serum of mice与正常组相比,模型组小鼠显著降低了TNF-α的分泌(P<0.05),说明环磷酰胺诱导下小鼠TNF-α的分泌受到了抑制。与模型组相比,AGS低、中、高剂量组的TNF-α分泌提升效果均显著(P<0.05),中剂量组与正常组小鼠无明显差异(P>0.05)。说明AGS能较好地恢复环磷酰胺诱导的TNF-α分泌降低的情况。
与正常组相比,模型组小鼠显著降低了IL-6的分泌(P<0.05),说明环磷酰胺诱导下小鼠IL-6的分泌受到了抑制。与模型组相比,AGS低、中、高剂量组的IL-6分泌都有所提升,具有剂量依赖性,且各剂量组小鼠提升IL-6分泌的效果与模型组差异显著(P<0.05),高剂量组与正常组小鼠无明显差异(P>0.05)。说明AGS能较好地恢复环磷酰胺诱导的IL-6分泌降低的情况。
与正常组相比,模型组小鼠显著降低了IFN-γ的分泌(P<0.05),说明环磷酰胺诱导下小鼠IFN-γ的分泌受到了抑制。与模型组相比,AGS低、中、高剂量组的IFN-γ分泌提升效果均显著(P<0.05)。中、高剂量组与正常组小鼠无明显差异(P>0.05)。说明AGS能较好地恢复环磷酰胺诱导的IFN-γ分泌降低的情况。
NF-κB信号通路通过胞外细胞因子与受体的结合,激活IκB激酶,进而调节炎症、癌症、自身免疫疾病等[40]。TNF-α能作为NF-κB经典通路的细胞因子,激活NF-κB信号通路。JAK-STAT信号通路主要由细胞外信号因子、酪氨酸激酶相关受体、JAK激酶和转录因子STAT四部分组成[41]。其中IL-6、INF-γ可作为细胞外信号因子,与细胞膜上相应的IL-6受体、INF-γ受体相结合,受体与JAK激酶相结合,实现信号从细胞外到细胞内的传递,从而参与免疫调节及细胞的增殖、分化和凋亡。AGS对TNF-α、IL-6、INF-γ分泌的提高或许提示着其免疫增强活性与NF-κB信号通路、JAK-STAT信号通路有关。
3. 结论
本论文通过对体外巨噬细胞RAW264.7和体内注射环磷酰胺建立免疫低下小鼠模型两方面探究AGS的免疫增强活性。结果表明AGS各浓度组均能促进体外巨噬细胞吞噬中性红大分子,提升TNF-α、IL-6、IFN-γ分泌的能力;AGS中剂量组和高剂量组均能缓解胸腺、脾脏萎缩和质量减轻现象,增强巨噬细胞的吞噬能力,提高巨噬细胞和脾细胞的增殖活性,增强NK细胞活性,提高CD4+/CD8+水平,提升小鼠分泌TNF-α、IL-6、IFN-γ的能力。综上所述,AGS可通过对免疫器官、免疫细胞和细胞因子等方面进行调节,从而起到免疫增强的作用。
-
![]()
![]()
图 2 AGS对小鼠巨噬细胞增殖活性的影响
Figure 2. Effects of AGS on proliferation of macrophages in mice
![]()
图 4 AGS对小鼠脾细胞中自然杀伤细胞(NK细胞)活力的影响
Figure 4. Effects of AGS on NK cell activity of splenocytes in mice
![]()
图 5 AGS对小鼠脾脏T淋巴细胞亚群的影响
注:A:正常组;B:模型组;C:低剂量组;D:中剂量组;E:高剂量组。
Figure 5. Effect of AGS on T lymphocyte subsets of spleen in mice
![]()
图 6 AGS对小鼠血清细胞因子的影响
注:A:TNF-α;B:IL-6;C:IFN-γ;*表示与正常组相比,差异显著(P<0.05);#表示与模型组相比,差异显著(P<0.05)。
Figure 6. Effects of AGS the release of cytokines in serum of mice
表 1 各组小鼠给药处理表
Table 1 Dosing and treatment of mice in each group
序号 组别 是否注射环磷酰胺 第11~20 d灌胃 1 正常组 否 去离子水 2 模型对照组 是 去离子水 3 低剂量组 是 0.5 mg/kg 4 中剂量组 是 5 mg/kg 5 高剂量组 是 20 mg/kg  下载: 导出CSV
下载: 导出CSV
表 2 AGS对RAW264.7吞噬能力的影响
Table 2 Effects of AGS on the phagocytosis of RAW264.7 cells
组别 吸光度 正常组 0.282±0.014 200(μg/mL) 0.329±0.018* 500(μg/mL) 0.321±0.006* 1000(μg/mL) 0.320±0.007* LPS 0.338±0.015* 注:*表示与正常组相比,差异显著(P<0.05)。
下载: 导出CSV
表 4 AGS对免疫低下小鼠碳廓清指数、吞噬指数的影响
Table 4 Effect of AGS on carbon clearance index and phagocytic index in immunocompromised mice
组别 碳廓清指数 吞噬指数 正常组 0.0689±0.0019 0.3551±0.0179 模型组 0.0199±0.0015* 0.0993±0.0119* 低剂量组 0.0210±0.0020* 0.1110±0.0116* 中剂量组 0.0339±0.0030*# 0.1888±0.0069*# 高剂量组 0.0409±0.0014*# 0.2489±0.0266*#
下载: 导出CSV
表 5 AGS对小鼠脾脏T淋巴细胞亚群的影响
Table 5 Effect of AGS on T lymphocyte subsets of spleen in mice
组别 CD4+(%) CD8+(%) CD4+/CD8+ 正常组 72.33±0.40 21.63±0.64 3.35±0.11 模型组 56.17±0.57* 35.80±0.26* 1.57±0.02* 低剂量组 58.10±1.21* 27.80±0.87*# 2.09±0.11*# 中剂量组 66.23±0.40*# 22.13±0.57# 2.99±0.09*# 高剂量组 58.43±0.32*# 29.50±0.79*# 1.98±0.04*#
下载: 导出CSV
-
[1] 姜会超, 刘爱英, 宋秀凯, 等. 重金属胁迫对刺参胚胎发育的影响[J]. 水生生物学报,2014,38(2):393−400. [JIANG H C, LIU A Y, SONG X K, et al. The toxic effects of heavy metals on the embryonic development of Apostichopus[J]. Acta Hydrobiologica Sinica,2014,38(2):393−400. doi: 10.7541/2014.56 [2] 段续, 王辉, 任广跃, 等. 海参的干制技术及其研究进展[J]. 食品工业科技,2012,33(10):427−431. [DUAN X, WANG H, REN G Y, et al. Research progress of dry-cure technology of sea cucumber[J]. Science and Technology of Food Industry,2012,33(10):427−431. doi: 10.13386/j.issn1002-0306.2012.10.103 [3] DAI Y L, KIM E A, LUO H M, et al. Characterization and anti-tumor activity of saponin-rich fractions of South Korean sea cucumbers (Apostichopus japonicus)[J]. International Journal of Food Science and Technology,2020,57(6):2283−2292. doi: 10.1007/s13197-020-04266-z
[4] WANG Z, ZHANG H, YUAN W, et al. Antifungal nortriterpene and triterpene glycosides from the sea cucumber Apostichopus japonicus selenka[J]. Food Chemistry,2012,132(1):295−300. doi: 10.1016/j.foodchem.2011.10.080
[5] 刘昕, 刘京熙, 张健, 等. 仿刺参卵多糖的分离纯化及体外抗肿瘤活性[J]. 食品科学,2016,37(23):105−110. [LIU X, LIU J X, ZHANG J, et al. Purification and antitumor activity in vitro of polysaccharides from Apostichopus japonicus spawn[J]. Food Science,2016,37(23):105−110. doi: 10.7506/spkx1002-6630-201623018 [6] 向怡卉, 苏秀榕, 董明敏, 等. 复合蛋白酶水解海参生殖腺工艺的研究[J]. 食品工业科技,2007,28(3):143−144, 146. [XIANG Y H, SU X R, DONG M M, et al. Study on hydrolysis of sea cucumber gonads by compound protease[J]. Science and Technology of Food Industry,2007,28(3):143−144, 146. doi: 10.3969/j.issn.1002-0306.2007.03.042 [7] 钱颖, 黄容容, 孙锐, 等. 人参皂苷Rh2对免疫低下小鼠的免疫调节作用[J]. 医药导报,2018,37(12):1446−1454. [QIAN Y, HUANG R R, SUN Y, et al. Effect of ginsenoside Rh2 on immune regulation of immunocompromised mice[J]. Herald of Medicine,2018,37(12):1446−1454. [8] 吴雨龙, 朱华, 张艺鏻, 等. 菊苣多糖对免疫抑制小鼠免疫功能的影响[J]. 食品工业科技,2021,42(3):284−289, 337. [WU Y L, ZHU H, ZHANG Y L, et al. Effect of chicory polysaccharide on immune function in immunosuppressed mice[J]. Science and Technology of Food Industry,2021,42(3):284−289, 337. doi: 10.13386/j.issn1002-0306.2020030168 [9] GE Y, LI C, REN H, et al. Effects of ginseng saponin Rh_(2) injection on immune function of H_(22) cancer mice[J]. Special Wild Economic Animal and Plant Research,2002,24(3):4−7.
[10] 张长城, 姜美杰, 赵海霞, 等. 竹节参总皂苷对环磷酰胺致免疫低下小鼠免疫功能的影响[J]. 中成药,2011,33(7):1134−1138. [ZHANG C C, JIANG M J, ZHAO H X, et al. Effects of total saponins of Panax japonicus rhizoma on cyclophosphamide-induced immunosuppressed mice[J]. Chinese Traditional Patent Medicine,2011,33(7):1134−1138. doi: 10.3969/j.issn.1001-1528.2011.07.011 [11] 王静凤, 傅佳, 王玉明, 等. 革皮氏海参皂苷对小鼠免疫功能的调节作用[J]. 中国海洋大学学报(自然科学版),2010,40(2):28−32. [WANG J F, FU J, WANG Y M, et al. Effects of saponins of Pearsonothuria graeffei on immune regulation in mice[J]. Periodical of Ocean University of China,2010,40(2):28−32. doi: 10.16441/j.cnki.hdxb.2010.02.005 [12] 翟星辰. 壳寡糖免疫增强及对肾癌抑制作用的研究[D]. 哈尔滨: 哈尔滨工业大学, 2019. ZHAI X C. Research on immune enhancement of chitosan oligosaccharides and its inhibitory effects against renal carcinoma[D]. Harbin: Harbin Institute of Technology, 2019.
[13] 董婧媛. 海参皂苷的提取工艺及功能研究[D]. 天津: 天津科技大学, 2020. DONG J Y. Study on extraction process and function of sea cucmber saponin[D]. Tianjing: Tianjin University of Science & Technology, 2020.
[14] YU J H, CONG L X, WANG C M, et al. Immunomodulatory effect of Schisandra polysaccharides in cyclophosphamide-induced immunocompromised mice[J]. Experimental and Therapeutic Medicine,2018,15:4755−4762.
[15] 刘仁杰, 王春凤, 王刚, 等. 林蛙油活性肽对小鼠免疫功能的影响[J]. 东北林业大学学报,2017,45(1):82−85, 89. [LIU R J, WANG C F, WANG G, et al. Effect of bioactive peptides from the oviductus ranae on immunologic function of mice[J]. Journal of Northeast Forestry University,2017,45(1):82−85, 89. doi: 10.3969/j.issn.1000-5382.2017.01.017 [16] 段炳南, 陈庆林. 绞股蓝总皂甙对小鼠腹腔巨噬细胞内酶活性及吞噬功能的影响[J]. 江西医学院学报,2007,47(3):38−40. [DUAN B N, CHEN Q L. Effects of gypenosides on enzyme activity and phagocytic capacity of peritoneal macrophage in mice[J]. Acta Academiae Medicinae Jiangxi,2007,47(3):38−40. [17] 王军. MIF, TGFβ, IFNγ基因多态性与脊柱结核易感性及在椎间盘中的表达与其临床资料的关联研究[D]. 南宁: 广西医科大学, 2017. WANG J. Relationship between MIF, TGFβ, IFNγ gene polymorphism and spinal tuberculosis suscepyibility and expression in intervertebral disc and its clinical data[D]. Guangxi: Guangxi Medical University, 2017.
[18] 张禹. 基于介孔羟基磷灰石与荧光碳点的免疫佐剂效应研究[D]. 广州: 暨南大学, 2017. ZHANG Y. The research on the immune adjuvant effects of mesoporous hydroxylapatite and fluorescent carbon dots[D]. Guangzhou: Jinan University, 2017.
[19] AMININ D L, AGAFONOVA I G, BERDYSHEV E V, et al. Immunomodulatory properties of cucumariosides from the edible far-eastern Holothurian Cucumaria japonica[J]. Journal of Medicinal Food,2001,4(3):127−135. doi: 10.1089/109662001753165701
[20] SAMOILOVA E B, HORTON J L, HILLIARD B, et al. IL-6-deficient mice are resistant to experimental autoimmune encephalomyelitis: Roles of IL-6 in the activation and differentiation of autoreactive T cells[J]. The Journal of Immunology,1998,161(12):6480−6486.
[21] WANG X, WANG Z Q, WU H H, et al. Sarcodon imbricatus polysaccharides protect against cyclophosphamide-induced immunosuppression via regulating Nrf2-mediated oxidative stress[J]. Accepted Manuscript,2018,120:736−744.
[22] BHARDWAJ J, CHAUDHARY N, SEO H J, et al. Immunomodulatory effect of tea saponin in immune T-cells and T-lymphoma cells via regulation of Th1, Th2 immune response and MAPK/ERK2 signaling pathway[J]. Immunopharmacology and Immunotoxicology,2014,36(3):202−210. doi: 10.3109/08923973.2014.909849
[23] SINGH K P, GUPTA R K, SHAU H, et al. Effect of ASTA-Z 7575 (INN Maphosphamide) on human lymphokine-activated killer cell induction[J]. Immunopharmacology and Immunotoxicology,1993,15(5):528−538.
[24] DENG J, ZHONG Y F, WU Y P, et al. Carnosine attenuates cyclophosphamide-induced bone marrow suppression by reducing oxidative DNA damage[J]. Redox Biology,2018,14:1−6. doi: 10.1016/j.redox.2017.08.003
[25] PRATHEESHKUMAR P, KUTTAN G. Ameliorative action of Vernonia cinerea L. on cyclophosphamide-induced immunosuppression and oxidative stress in mice[J]. Inflammopharmacology,2010,18(4):197−207. doi: 10.1007/s10787-010-0042-8
[26] 王飞. 饲养环境中硫化氢暴露引起小鼠免疫功能变化的初步调查[D]. 保定: 河北农业大学, 2019. WANG F. Preliminary investigation on the changes of immune function in mice induced by hydrogen sulfide exposure in breeding environment[D]. Baoding: Hebei Agricultural University, 2019.
[27] 杨杰, 卫东锋, 王文潇, 等. 五指毛桃水提物对免疫抑制小鼠细胞免疫的影响[J]. 中药药理与临床,2015,31(6):111−114. [YANG J, WEI D F, WANG W X, et al. Effects of aqueous extract of Ficus hirta on cellular immunity in immunosuppressed mice[J]. Pharmacology and Clinics of Chinese Materia Medica,2015,31(6):111−114. doi: 10.13412/j.cnki.zyyl.2015.06.033 [28] DUGGINA P, KALLA C M, VARIKASUVU S R, et al. Protective effect of centella triterpene saponins against cyclophosphamide-induced immune and hepatic system dysfunction in rats: Its possible mechanisms of action[J]. Journal of Physiology and Biochemistry,2015,71:435−454. doi: 10.1007/s13105-015-0423-y
[29] 杜双双. 蚕丝蛋白肽免疫调节及与化疗的联合作用[D]. 天津: 天津医科大学, 2018. DU S S. The immunomodulation and combinedeffect with chemotherapy of silk fibroinpeptide[D]. Tianjin: Tianjin Medical University, 2018.
[30] CHEN L X, QI Y L, QI Z, et al. A comparative study on the effects of different parts of Panax ginseng on the immune activity of cyclophosphamide-induced immunosuppressed mice[J]. Molecules,2019,24(6):1096. doi: 10.3390/molecules24061096
[31] YU Q, NIE S P, LI W J, et al. Macrophage immunomodulatory activity of a purified polysaccharide isolated from Ganoderma atrum[J]. Phytotherapy Research,2013,27(2):186−191. doi: 10.1002/ptr.4698
[32] LEIRO J M, CASTRO R, ARRANZ J A, et al. Immunomodulating activities of acidic sulphated polysaccharides obtained from the seaweed Ulva rigida C. Agardh[J]. International Immunopharmacology,2007,7(7):879−888. doi: 10.1016/j.intimp.2007.02.007
[33] 苗明三, 刘会丽, 杨亚蕾, 等. 无花果多糖对免疫抑制小鼠腹腔巨噬细胞产生IL-1α、脾细胞体外增殖、脾细胞产生IL-2及其受体的影响[J]. 中国现代应用药学,2009,26(7):525−528. [MIAO M S, LIU H L, YANG Y L, et al. Effect of Ficus carica oolysaccharide on the levels of interleukin-la produced by peritoneal macro-phages, lymphocyte proliferation, interleukin-2 and its receptor produced by lymphocyte in immu-nosuppressive mice[J]. Chinese Journal of Modern Applied Pharmacy,2009,26(7):525−528. [34] ROSISA M, ALEKSANDAR S, DENITSA A, et al. In vitro antitumour and immunomodulating activity of saponins from Astragalus glycyphyllos[J]. Biotechnology & Biotechnological Equipment,2022,35(1):1948−1955.
[35] 董毅, 李瑞. 运动与自然杀伤细胞抗病毒等功能的关系和机制[J]. 中国体育科技,2020,56(5):3−13. [DONG Y, LI R. The relationship and underlying mechanism between exercise and functions of NK Cells[J]. China Sport Science and Technology,2020,56(5):3−13. [36] 王静凤, 王奕, 赵林, 等. 日本刺参的抗肿瘤及免疫调节作用研究[J]. 中国海洋大学学报(自然科学版),2007,37(1):93−96,102. [WANG J F, WANG Y, ZHAO L, et al. Effects of Apostichopus japonicus on antitumor and immune regulation in S180 bearing mice[J]. Periodical of Ocean University of China,2007,37(1):93−96,102. [37] 白军, 李博文, 刘淑红. 金丝桃苷对小鼠T淋巴细胞亚群及血清细胞因子的影响[J]. 动物医学进展,2017,38(6):48−51. [BAI J, LI B W, LIU S H. Effects of hyperoside on T cell subset and serum cytokines in mice[J]. Progress in Veterinary Medicine,2017,38(6):48−51. doi: 10.3969/j.issn.1007-5038.2017.06.011 [38] 曹晓军, 倪慧萍. 传染性单核细胞增多症患儿T细胞亚群变化及临床意义[J]. 江苏大学学报(医学版),2007,17(5):429−431. [CAO X J, NI H P. Change of T cell subsets in children with infectious mononucleosis and its clinical meanings[J]. Journal of Jiangsu University (Medicine Edition),2007,17(5):429−431. doi: 10.13312/j.issn.1671-7783.2007.05.016 [39] 赵建国, 刘玲艳, 朱颖越, 等. 合欢皮总皂苷急性毒理学研究[J]. 天然产物研究与开发,2010,22(4):582−586. [ZHAO J G, LIU L Y, ZHU Y Y, et al. Research on emergency toxicology of total saponin in silktree Albizia bark[J]. Natural Product Reseaech and Development,2010,22(4):582−586. doi: 10.3969/j.issn.1001-6880.2010.04.010 [40] 焦园园, 王萍, 赵凤, 等. 基于氧化应激探究PCI损伤及中医药防治进展[J]. 中国实验方剂学杂志,2020,26(4):214−225. [JIAO Y Y, WANG P, ZHAO F, et al. Research on PCI damage based on oxidative stress and progress in prevention and treatment of traditional Chinese medicine[J]. Chinese Journal of Experimental Traditional Medical Formulae,2020,26(4):214−225. doi: 10.13422/j.cnki.syfjx.20200439 [41] 唐爱存, 王明刚, 卢秋玉, 等. 葫芦茶苷调控JAK/STAT信号通路抗乙肝病毒作用及其机制研究[J]. 中药药理与临床,2017,33(1):74−77. [TANG A C, WANG M G, LU Q Y, et al. Study on anti-hepatitis B virus activities and mechanisms of tadehaginoside by regulating JAK/STAT signaling pathway[J]. Pharmacology and Clinics of Chinese Materia Medica,2017,33(1):74−77. doi: 10.13412/j.cnki.zyyl.2017.01.021
 下载:
下载:











计量
- 文章访问数: 174
- HTML全文浏览量: 37
- PDF下载量: 9
